

¿Cómo se adaptan los organismos al cambio climático?
Las inversiones cromosómicas de Drosophila subobscura: el caso de las poblaciones de Serbia

Es conocido el hecho de que las inversiones cromosómicas de las poblaciones naturales de la especie Drosophila subobscura son adaptativas con respecto a las variaciones del ambiente. Se ha observado que las inversiones cambian en frecuencia a lo largo del tiempo según la predicción del calentamiento global de nuestro planeta. Estos resultados se ponen de manifiesto en nuestros estudios realizados a poblaciones serbias de D. subobscura. Así, el polimorfismo cromosómico por inversiones cambia según las estaciones del año y también a largo plazo (períodos de 10-15 años) de acuerdo con lo que se esperaría según el calentamiento global. Por tanto, las inversiones cromosómicas de esta especie son unos buenos indicadores del cambio climático y también son útiles para estudiar cómo se pueden adaptar los organismos.
Palabras clave: Drosophila subobscura, inversión cromosómica, adaptación, selección natural, cambio climático.
La genética y el cambio climático
Los seres vivos tienen la capacidad de adaptarse a los cambios ambientales. Las adaptaciones no son más que unas características del diseño de los organismos que les permiten sobrevivir y reproducirse. Esta capacidad adaptativa, para ser útil, tiene que poder pasar a la descendencia, por tanto tiene que ser hereditaria. Es por esta razón que la genética es tan importante a la hora de explicar cómo afrontan las especies el reto de un cambio ambiental tan intenso y general como es el climático. Actualmente sabemos que los genomas son unos grandes reservorios de variabilidad genética que está a disposición de los organismos para poder llevar a cabo las adaptaciones y no desaparecer.
«Las adaptaciones son unas características del diseño de los organismos que les permiten sobrevivir y reproducirse»
El cambio climático actual representa una gran oportunidad para la investigación, porque podemos aprender mucho sobre los mecanismos genéticos de la adaptación de las especies. Estos estudios se pueden hacer recogiendo muestras de la misma especie a lo largo del tiempo o en diferentes lugares geográficos (más fríos o más cálidos, por ejemplo) y ver qué diferencias se encuentran para algunos genes o marcadores genéticos. Con la información de estas observaciones hechas en la naturaleza se puede intentar realizar experimentos en el laboratorio simulando condiciones climáticas particulares (normalmente variando la temperatura) y analizar si los organismos reaccionan igual con respecto a los mismos genes o marcadores genéticos. Estos resultados pueden darnos información sobre la base genética de los mecanismos adaptativos, en este caso, con respecto al cambio climático. Actualmente, y gracias a las llamadas técnicas del «ADN antiguo» podemos incluso analizar fragmentos del genoma de organismos fósiles, tanto de especies extintas como actuales, y analizar cuáles de estos trozos del genoma pueden ser clave en las adaptaciones climáticas. Tampoco podemos olvidar que a veces la adaptación de los organismos no pasa porque un gen tenga una variante u otra, sino por los mecanismos de regulación de estos genes. La información con respecto a la expresión génica es muy valiosa porque nos podría ayudar a prever la adaptación o no de poblaciones de organismos frente a los cambios ambientales producidos por el calentamiento global.

Por otro lado, la genética nos puede ser útil en el otro sentido, en una vertiente más práctica. Si conocemos qué individuos (o poblaciones) presentan unas variantes genéticas que les permiten sobrevivir mejor en unas determinadas condiciones ambientales se pueden reconstruir algunos ecosistemas muy degradados por el cambio climático. Esta aproximación se ha aplicado a la reconstrucción de algunos ecosistemas de los Estados Unidos (Wymore et al., 2014). Esta técnica pasa por el estudio genético y ecológico de muchos organismos, desde las bacterias hasta las plantas y los animales.
En este trabajo pretendemos mostrar nuestro estudio sobre los mecanismos genéticos de las especies que les permiten la adaptación, concretamente al cambio climático. Nuestra especie es un tipo de mosca de la fruta, la llamada Drosophila subobscura, y los marcadores genéticos que estudiamos son las inversiones cromosómicas, es decir, fragmentos de cromosomas que están situados en una orientación opuesta a la habitual.
La especie: ‘Drosophila subobscura’
Drosophila subobscura es un díptero muy utilizado en estudios de genética, pero sobre todo de evolución (figura 1). Tiene una distribución biogeográfica (Krimbas, 1993) que alcanza gran parte de Europa, excepto la parte central y norte de Escandinavia e Islandia. Al norte de África se encuentra hasta el desierto del Sáhara y en el Oriente Medio está presente hasta Irán. La zona que ocupa en los territorios de la antigua Unión Soviética es poco conocida. También se la puede encontrar en las islas del Mediterráneo, así como en las Azores, Madeira y Canarias. En 1978 se describió su presencia por primera vez en Chile (Mestres, Balanyà, Prevosti y Serra, 1993), y se expandió rápidamente por toda la región al oeste de los Andes, desde La Serena hasta Punta Arenas, a pesar de que se cree que en la zona sur de Chile su presencia podría ser estacional. Esta colonización seguramente provendría de una población mediterránea, pero aún no se conoce con exactitud el origen. Unos años después, en concreto en 1982, se encontró por primera vez en la costa oeste de Norteamérica, desde el sur de California hasta la región sur del estado canadiense de British Columbia. Hay que mencionar también que un pequeño núcleo colonizador se detectó alrededor de la ciudad argentina de Mar del Plata. Se cree que sería una colonización secundaria a partir de la principal de Chile. Hay una constante en las tres regiones ocupadas actualmente por la especie, todas ellas presentan la misma alternancia de climas: maritimooccidental y mediterráneo (hasta la zona semiárida predesértica).
«Una inversión es un fragmento de cromosoma que pasa a colocarse en el orden opuesto al normal. No representa ningún problema para los individuos que las presentan»
Una de las principales características de D. subobscura es que solo tiene seis cromosomas politénicos, que se llaman A (X o sexual), J, U, E, O y dot (puntiforme). Todos, a excepción del puntiforme, presentan inversiones cromosómicas (figura 2). Como ya se ha comentado, una inversión no es más que un fragmento de cromosoma que pasa a colocarse en el orden opuesto al normal. En D. subobscura las inversiones se denominan por una letra (que indica el cromosoma) y un número (que identifica la inversión concreta) o el símbolo «st» (estándar o referencia). Muchas veces un mismo cromosoma tiene varias inversiones y forman lo que se llama un «ordenamiento». Las diferentes inversiones que forman los ordenamientos se identifican por sus números separados por el signo «+». Finalmente, para mostrar si las inversiones están totalmente o parcialmente solapadas se utiliza el subrayado (por ejemplo O3+4). El hecho de tener inversiones no representa ningún problema para los individuos que las presentan, porque tienen todos los mismos genes, lo único es que están cambiados de orden. En cambio este hecho tiene una importancia evolutiva crucial, ya que los genes localizados dentro de la inversión tenderán a heredarse juntos. Si la combinación génica que contiene una inversión es favorable en el sentido de que le permite adaptarse mejor al medio ambiente donde vive (es decir, sobrevivir y reproducirse), esta será favorecida por la selección natural y por tanto aumentará en frecuencia a lo largo de las generaciones. Por esta razón, D. subobscura ha tenido tanto éxito como especie modelo en estudios evolutivos.
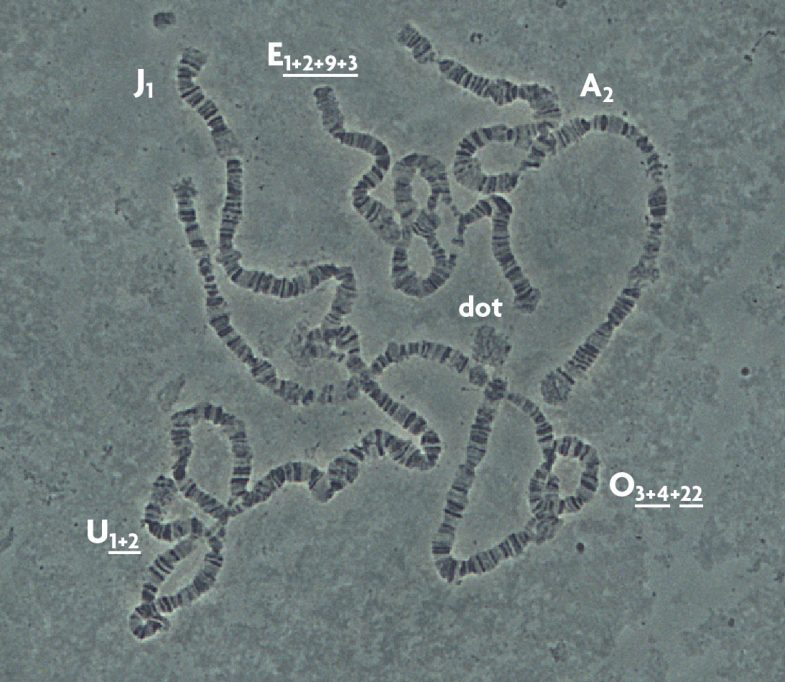
A partir de los años cincuenta del siglo pasado diferentes escuelas europeas de investigadores empezaron a analizar el contenido de las poblaciones naturales de D. subobscura con respecto a las inversiones. Estas presentaban variaciones entre diferentes poblaciones e incluso entre diferentes muestras de una misma localidad. Lo que más sorprendió a aquellos científicos era que las frecuencias de algunas de ellas cambiaban con la latitud, es decir, presentaban lo que se llama un cambio clinal. De esta manera, se observaba que unas inversiones eran muy abundantes en el sur de Europa y su frecuencia iba disminuyendo al muestrear más hacia el norte (como por ejemplo O3+4). En cambio, otras que eran poco abundantes al sur, iban aumentando en frecuencia al analizar poblaciones hacia el norte (como el caso de Ost). Hay dos explicaciones del fenómeno: una posibilidad sería que se tratase de un proceso histórico. En un cierto momento aparecería una inversión (hay diferentes mecanismos que las generan, pero no los explicaremos en este trabajo) y podría llegar a ser muy abundante en la población donde aparece. Con el movimiento de las drosófilas, la inversión se iría extendiendo pasivamente por Europa mostrando un gradiente, y evidentemente, cuanto más lejos de su origen se esperaría encontrarla con menor frecuencia. La segunda hipótesis se basaría en el valor adaptativo de las inversiones. Una inversión nueva, si aporta ventaja a los individuos que la presentan, aumentará en frecuencia gracias a la selección natural. Si esta ventaja es una adaptación a las condiciones climáticas (fundamentalmente temperatura y otros factores ligados a este parámetro), esperaremos que su abundancia dependa de la posición de las poblaciones a lo largo de un eje norte-sur. Durante muchos años se discutió cuál de las dos hipótesis explicaba la distribución clinal de muchas de las inversiones de la especie (Krimbas y Loukas, 1980). El hecho de encontrar en las dos regiones colonizadas del continente americano (Chile y la costa oeste de los Estados Unidos) clinas latitudinales para casi las mismas inversiones y con el mismo sentido dio un gran apoyo al valor adaptativo de las inversiones (Prevosti et al., 1988).
El cambio climático y las inversiones cromosómicas
Hay medidas meteorológicas que indican que la temperatura en nuestro planeta va aumentando. Por ejemplo parece que el calentamiento global es de 0,6 ºC en las tres décadas pasadas y de 0,8 ºC con respecto al siglo pasado (Hansen et al., 2006). También la nieve y el hielo parece que han disminuido cerca de un 10 % en nuestro planeta. En cambio, como consecuencia, el patrón de precipitaciones ha devenido irregular. De este hecho se resienten los seres vivos y lo ecosistemas de los que forman parte. Por ejemplo, se ha modificado el patrón de actividad estacional en muchos organismos, así como también el tiempo de floración, de la caída de las hojas o de las migraciones.
«Si la combinación génica que contiene una inversión es favorable porque le permite adaptarse mejor al medio ambiente donde vive, esta será favorecida por la selección natural»
Uno de los primeros investigadores en percatarse de la importancia del cambio de las frecuencias de las inversiones en el tiempo fue el doctor Antoni Prevosti. Así, en uno de sus trabajos (De Frutos y Prevosti, 1984) se dio cuenta tanto de la presencia de cambios estacionales como a largo plazo. Años después, en la última tesis que dirigió, relacionó los cambios a largo plazo en las inversiones cromosómicas con las variaciones en la temperatura (Orengo y Prevosti, 1996). Una vez más, Prevosti volvía a demostrar los efectos rápidos de la selección natural en D. subobscura. Estos trabajos tuvieron continuación de la mano del doctor Lluís Serra, que analizó el cambio a largo plazo de las inversiones cromosómicas en esta especie comparando los datos antiguos con nuevas capturas para una colección amplía de poblaciones europeas (un total de trece en un intervalo de quince años como mínimo) y también americanas, tanto del norte como del sur (trece poblaciones analizadas en un intervalo entre dieciocho y veinte años). Si el calentamiento era global, en las poblaciones europeas, pero también en las americanas, pasados unos años del origen de la doble colonización, las inversiones tendrían que cambiar en frecuencia según las expectativas del cambio climático. En general, las poblaciones analizadas tanto en Europa como en América presentaban diferencias a largo plazo tanto en el tipo de inversiones como en sus frecuencias, de manera que las inversiones consideradas «cálidas» iban ganando terreno con los tiempos (Balanyà, Huey, Gilchrist y Serra, 2009).

Los estudios de las inversiones cromosómicas en Serbia
¿Por qué era interesante averiguar qué pasaba en Serbia? Esta región biogeográfica era importante por diferentes razones. La mayoría de clinas latitudinales que históricamente se han estudiado en D. subobscura van en dirección sudoeste/nordeste, arrancando de la península Ibérica. Además, aunque no se puede afirmar, existe toda una serie de pruebas genéticas que hacen pensar que el origen de la colonización de América tuvo lugar a partir de alguna población del Mediterráneo occidental. Por tanto, era interesante ver si también las inversiones cambiaban y lo hacían en el sentido predicho por el calentamiento global en otra región europea diferente, en la que no se había llevado a cabo ningún estudio de clinas latitudinales ni de variación a lo largo de los años. Las poblaciones de D. subobscura de los Balcanes tienen un contenido en inversiones cromosómicas que se puede calificar de mediterráneo, pero es bastante diferente en algunos aspectos (tipo y frecuencia de algunas inversiones) del que se encuentra en la cuenca occidental (Krimbas, 1993). Serbia, como forma parte de los Balcanes, tiene como ventaja ser considerada un refugio por muchas especies durante la última glaciación. Este hecho la hace evolutivamente atractiva, ya que podría haber acumulado bastante variabilidad genética. Las poblaciones de D. subobscura de este país han sido muy estudiadas en materia de inversiones, gracias a la escuela fundada por el doctor Dragoslav Marinkovic, y por tanto se tenía mucha información de años atrás de este marcador genético (para una buena revisión se puede consultar Krimbas, 1993 y Zivanovic, Andjelkovic y Marinkovic, 2002).

El primer estudio de la serie lo realizamos a partir de unas muestras de la población de Avala, localizada a 18 km de Belgrado (figura 3) y situada a unos 450 metros sobre el nivel del mar (Zivanovic y Mestres, 2010a). Se hicieron capturas durante los meses de septiembre de 2003, 2004 y 2005. Si se comparaban con las muestras obtenidas en otras poblaciones serbias, parecía que las frecuencias de las inversiones consideradas «frías» (en concreto Jst, Ust, Este y Ost) habían disminuido, mientras que las de algunas calificadas de «cálidas» habían aumentado (J1, U1+2, U1+2+6, E1+2+9 y O3+4). También se observó alguna pequeña variación entre años consecutivos. Eso también es lógico, porque las condiciones ambientales de los meses anteriores a la captura condicionan los resultados con respecto a las inversiones cromosómicas. Hay que recordar también que muchas de las inversiones son adaptativas, no solamente con respecto a la temperatura, sino a otros factores, que pueden estar relacionados o no con esta. En otro trabajo, directamente relacionado con el anterior, se estudiaron los cambios con respecto a las estaciones (Zivanovic y Mestres, 2010b). En concreto se compararon las inversiones obtenidas en el mismo lugar (Avala) pero en una muestra de junio de 2004 con respecto a otra de septiembre del mismo año. Se encontraron cambios por algunas inversiones (localizadas en los cromosomas J, E y U) y en el mismo sentido que los observados por otros investigadores.

Estos descubrimientos nos animaron a continuar en esta línea de investigación, y para el siguiente estudio se escogió la población de Apatin, una zona húmeda del banco izquierdo del río Danubio (Zivanovic y Mestres, 2011). Se trata de un hábitat bastante estable y poco afectado por la actividad humana, idóneo para la investigación propuesta. Se tenían datos de los cromosomas analizados en 1994 y se hicieron nuevas capturas en 2008 y 2009. Una primera aproximación para ver si las inversiones cromosómicas varían con respecto a lo que sería esperable según el cambio climático es, como ya hemos explicado, estudiar si las inversiones consideradas como «cálidas» y como «frías», respectivamente, aumentan y disminuyen. Pero para ser precisos hay que estudiar si la temperatura cambia con los años. Por suerte, pudimos obtener datos de las temperaturas máxima, mínima y media, así como de la pluviosidad de la población, gracias a los registros del Servicio Hidrometeorológico de la República de Serbia. Las temperaturas mostraron una tendencia a aumentar, mientras que el régimen de lluvias era irregular (hecho también característico del cambio climático). Con respecto a la composición en inversiones, esta había variado en el tiempo, detectándose por primera vez en 2009 el ordenamiento U1+8+2, considerado «cálido». También presentaron un aumento significativo las inversiones «cálidas» U1+2, O3+4 y O3+4+22.
Posteriormente, se pensó en estudiar los cromosomas de otra población pero con respecto a cambios mensuales (período corto de tiempo) y también en relación a muestras de hacía quince años (largo plazo). La población serbia escogida fue Petnica (captura realizada el año 2010), a unos 100 km al sur de Belgrado (Zivanovic, Arenas y Mestres, 2012). El lugar de captura era un bosque de Carpinus betula. En este trabajo también se quiso hacer un estudio comparado mediante métodos de estadística multivariante de las poblaciones serbias de las que se tenían datos por las inversiones cromosómicas (un total de dieciocho muestras). Existían cambios mensuales en la composición cromosómica de las muestras. Eso puede ser debido a que la selección natural actuase en respuesta a los cambios ambientales que se van produciendo, pero también al hecho de que los individuos se pueden desplazar y pueden buscar los lugares por los que tengan una preferencia térmica. En cuanto a los estudios a largo plazo, en la muestra de 2010 se detectaron por primera vez inversiones «cálidas» (U1+8+2, E1+2+9+12 y O3+4+8) no observadas en la muestra de 1995. Una vez más se observó el aumento de las inversiones «cálidas» (A2, J1, U1+2 y O3+4) y la disminución de las «frías» (A2, Jst, Ust, Est y Ost). Un punto que hay que destacar del trabajo es que en Petnica se encontró un aumento de todas las temperaturas estudiadas (máxima, mínima y media), siendo significativo el cambio en la temperatura mínima.
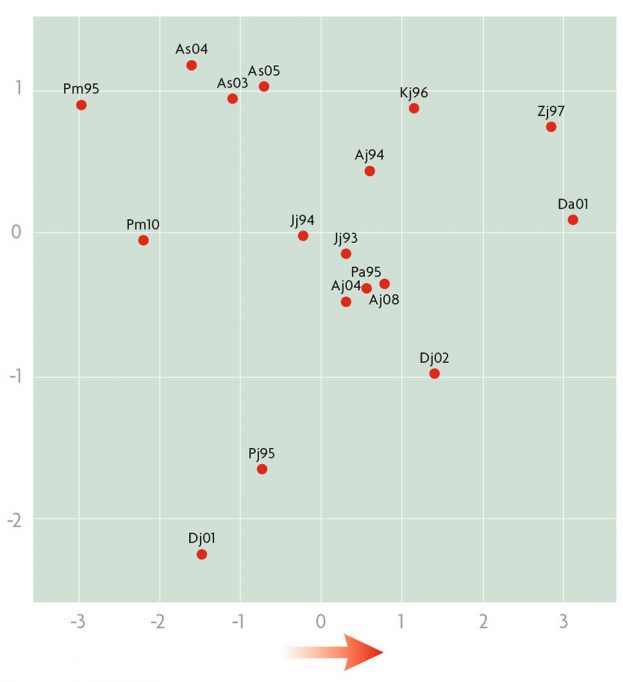
En cuanto al estudio de las inversiones en el conjunto de las poblaciones balcánicas, el análisis clúster agrupaba en general todas las «antiguas» en un grupo y todas las «recientes» en otro. Por tanto, este marcador genético se demostraba de nuevo que variaba en el tiempo. Al estudio de componentes principales, utilizando los datos de las inversiones y de las características climáticas de las poblaciones, la primera y la segunda componente explicaban respectivamente el 67,27 % y el 23,17 % de la variación total (figura 4). Lo que es interesante es que la primera explica las diferencias entre poblaciones debidas a temperatura y lluvias, es decir, desplazándose con respecto a ella en sentido positivo cada vez se encuentran poblaciones más cálidas y más secas. Por tanto, la composición en inversiones de las poblaciones es una respuesta de la selección natural a los cambios climáticos. Es más, este cambio sigue el sentido esperado por el efecto del calentamiento global del planeta. Este ámbito de estudio es fascinante y nuestra intención es continuar haciendo nuevos experimentos y análisis, como por ejemplo ver el efecto de las combinaciones de inversiones en una misma pareja de homólogos (cariotipos) o las interacciones entre inversiones de cromosomas de parejas diferentes. Nuestros estudios preliminares parecen indicar que las inversiones, por ellas mismas, tienen un valor adaptativo más importante que el de los cariotipos (Zivanovic, Arenas y Mestres, 2014). En resumen, las inversiones cromosómicas son un buen marcador del cambio climático y, además, permiten estudiar cómo las especies se pueden adaptar.
REFERENCIAS
Balanyà, J., Huey, R. B., Gilchrist, G. W., & Serra, L. (2009). The chromosomal polymorphism of Drosophila subobscura: A microevolutionary weapon to monitor global change. Heredity, 103, 364–367. doi: 10.1038/hdy.2009.86
De Frutos, R., & Prevosti, A. (1984). Temporal changes of chromosomal polymorphism in natural populations of Drosophila subobscura. Genetica, 63, 181–187. doi: 10.1007/BF00128411
Hansen, J., Sato, M., Ruedy, R., Lo, K., Lea, D. W., & Medina-Elizade, M. (2006). Global temperature change. Proceedings National Academy of Sciences, 103, 14288–14293. doi: 10.1073/pnas.0606291103
Krimbas, C. B. (1993). Drosophila subobscura: Biology, genetics and inversion polymorphism. Hamburgo: Verlag Dr Kovac.
Krimbas, C. B., & Loukas, M. (1980). The inversion polymorphism of Drosophila subobscura. Evolutionary Biology, 12, 163–234.
Mestres, F., Balanyà, J., Prevosti, A., & Serra, L. (1993). Genética evolutiva de la especie colonizadora Drosophila subobscura. Mundo Científico, 13, 408–416.
Orengo, D. J., & Prevosti, A. (1996). Temporal changes in chromosomal polymorphism of Drosophila subobscura related to climatic changes. Evolution, 50, 1346–1350. doi: 10.2307/2410676
Prevosti, A., Ribo, G., Serra, L., Aguade, M., Balaña, J., Monlcus, M., & Mestres, F. (1988). Colonization of America by Drosophila subobscura: Experimental in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proceedings National Academy of Sciences, 85, 5597–5600. doi: 10.1073/pnas.85.15.5597
Wymore, A. S., Bothwell, H. M., Compson, Z. G., Lamit, L. J., Walker, F. M., Woolbright S. A., & Witham T. G. (2014). Community genetics applications for forest biodiversity and policy: Planning for the future. In T. Fenning (Ed.). Challenges and opportunities for the world’s forest in the 21st century (pp. 707–726) Londres: Springer.
Zivanovic, G., Andjelkovic, M. & Marinkovic, D. (2002). Chromosomal inversion polymorphism of Drosophila subobscura from South-eastern part of Europe. Journal of Zoological Systematics and Evolutionary Research, 40(4), 201–204. doi: 10.1046/j.1439-0469.2002.00189.x
Zivanovic, G., & Mestres, F. (2010a). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. I. Analysis over several years. Hereditas, 147, 70–81. doi: 10.1111/j.1601-5223.2009.02163.x
Zivanovic, G. & Mestres, F. (2010b). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. II. Seasonal component analysis. Hereditas, 147, 82–89. doi: 10.1111/j.1601-5223.2010.02164.x
Zivanovic, G., & Mestres, F. (2011). Changes in chromosomal polymorphism and global warming: The case of Drosophila subobscura from Apatin (Serbia). Genetics and Molecular Biology, 34, 489–495. doi: 10.1590/S1415-47572011000300020
Zivanovic, G., Arenas, C., & Mestres, F. (2012). Short- and long-term changes in chromosomal inversion polymorphism and global warming: Drosophila subobscura from the Balkans». Israel Journal of Ecology and Evolution, 58, 289–311. doi: 10.1560/IJEE.58.4.289
Zivanovic, G., Arenas, C., & Mestres, F. (2014). Inversion polymorphism in two serbian natural populations of Drosophila subobscura: Analysis of long-term changes. Russian Journal of Genetics, 50, 638–644. doi: 10.7868/S0016675814060150




