

Historia y realidad del género ‘Homo’
¿Qué es y por qué lo consideramos como tal?
Los paleoantropólogos que se ocupan de cómo la naturaleza se organiza en especies y cómo deberíamos denominarlas suelen ser acusados de «limitarse a discutir sobre nombres». Esta idea implica que la taxonomía básica es una tarea administrativa aburrida que se debería solucionar rápidamente, o incluso ignorar, para poder llegar cuanto antes a las cuestiones realmente interesantes sobre la evolución humana. Sin embargo, la realidad es que nunca entenderemos la trama de esa intrincada obra que es la evolución humana si no identificamos con precisión a los actores que participaron en ella. Este artículo revisa la formación del concepto actual de género Homo, que resulta sumamente vago, como trasfondo para instar a realizar una definición más racional.
Palabras clave: taxonomía, géneros, género Homo, paleoantropología.
El género ‘Homo’: Enfoque de la cuestión
La primera cuestión que surge al definir el género Homo es la naturaleza de la propia unidad. Hoy en día, se acepta mayoritariamente que en principio los géneros no deberían ser solamente monofiléticos, sino holofiléticos. Es decir, un género debería contener todas las especies descendientes de un ancestro común concreto, y ninguna otra. Pero no existe una regla, ni siquiera una empírica, que diga cuántas especies debería haber en cada género, o lo remoto que puede llegar a ser ese ancestro común. Algunos géneros actuales bien establecidos son ricos en especies, mientras que otros tienen muchas menos y algunos incluyen una única especie. En parte, esta variedad se da porque algunos géneros parecen más intrínsecamente dispuestos que otros a la diversificación; pero en parte también se debe a una flexibilidad taxonómica que reconoce que no existen fronteras «naturales» para un género (como sí las hay, al menos en teoría, para las especies).
«Un género debería contener toda las especies descendientes de un antepasado común concreto, y ninguna otra»
Afortunadamente, en el mundo de los seres vivos, los géneros tienden a ser fácilmente reconocibles como unidades Gestalt.1 Esto se debe a que las distancias morfológicas entre géneros relacionados suelen ser mucho más grandes que los que separan a especies del mismo género, puesto que a medida que se expande la red, las diferencias se acumulan de forma natural, aunque a un ritmo irregular. Los juicios taxonómicos son inherentemente más complicados utilizando registros fósiles que seres vivos, no solo porque la información disponible es necesariamente más limitada, sino también porque las pruebas que sí tenemos tienden a ser más bien incompletas y están dispersas en el tiempo y el espacio.
Nuestra comprensión actual de cada rama de la ciencia está profundamente condicionada por la historia particular de cada campo. Y en el caso de la paleoantropología es especialmente cierto que la creencia actual es producto – hasta un punto alarmante– de la secuencia completamente arbitraria en la que se descubrieron los componentes del registro fósil conocido.
Nuestras ideas paleoantropológicas actuales también se han visto profundamente afectadas en el último par de siglos por las enormes fluctuaciones en nuestras ideas sobre lo que son y deberían ser los géneros. Debido a esta realidad, estas consideraciones sobre el género Homo son principalmente históricas. Porque casi con toda seguridad, si mañana se descubriera el registro fósil homínido al completo en lugar de ir encontrando elementos poco a poco a lo largo de siglo y medio, lo veríamos de forma muy distinta a como lo vemos hoy.
Primeras nociones del género ‘Homo’
El concepto original del género Homo, acuñado por Linneo (Linnaeus, 1758), se podía comparar estrechamente con la superfamilia Hominoidea actual, puesto que abarcaba no solo a Homo sapiens sino también a Homo troglodytes, una especie que incluía tanto a chimpancés como a orangutanes. La noción de que Homo sapiens podría haber compartido género con parientes extintos hoy en día no apareció hasta 1864, cuando William King clasificó como Homo neanderthalensis un esqueleto fósil parcial claramente antiguo que se había encontrado en el valle (Thal en alemán) de Neander en Alemania (King, 1864). Durante tres décadas, no obstante, no se describió ninguna nueva forma extinta de Homo, principalmente porque la discusión sobre los orígenes biológicos de la humanidad continuó en ausencia de cualquier fósil, aparte de unos pocos que se asemejaban a los neandertales. Aunque no todos lo veían como un obstáculo.
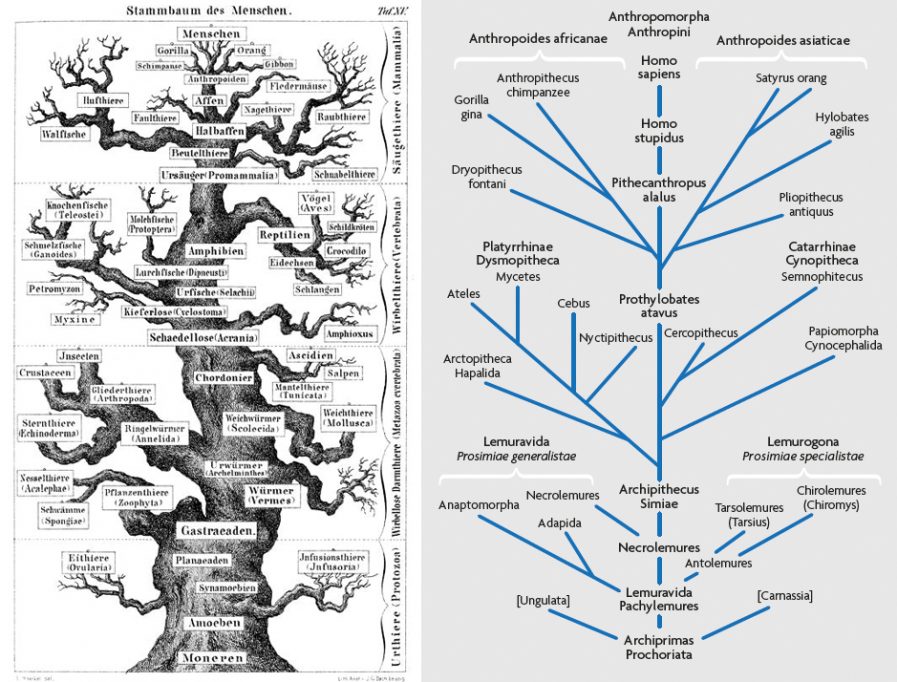
La Figura 1 muestra la noción de Ernst Haeckel del árbol de la vida, publicada en 1868, cuando, aparte de los neandertales, apenas existía un registro fósil humano. El dibujo de Haeckel representa la clásica imagen de un árbol con un tronco estrecho y nudoso y Homo sapiens (Menschen) en la cima. En una filogenia de los primates posterior y más detallada (Figura 2), Haeckel fue incluso más lejos, aunque en aquel momento él solo podía imaginar lo que la historia fósil de la humanidad revelaría. Pero sabía que debían existir conexiones anteriores entre los seres humanos y el resto de la naturaleza, por lo que insertó en su filogenia dos especies hipotéticas, Homo stupidus (que no el neandertal, si es lo que usted estaba pensando) y Pithecanthropus alalus, conectados por ascendencia y descendencia directa y lineal (Haeckel, 1868).
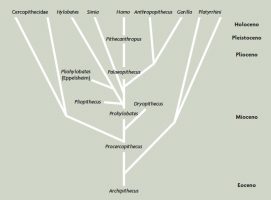
Por eso en 1891, cuando Eugene Dubois descubrió en Trinil (Java) la calota craneal 2 de la especie fósil de homínidos que más tarde se conocería como Pithecanthropus erectus, le pareció perfectamente lógico considerar al espécimen como un antepasado lineal de Homo sapiens, ya que tenía un cerebro pequeño pero claramente humano en cierto sentido. Este increíble hallazgo parecía encajar a la perfección en el constructo de Haeckel; hasta tal punto que Dubois utilizó para su fósil el nombre que Haeckel daba al género, Pithecanthropus (“simio-hombre”). Esta posición intermedia se refleja en el diagrama filogenético de Dubois de 1896 (Figura 3, véase Theunissen, 1988). Gracias a los esfuerzos de Dubois, pero también de muchos otros académicos influyentes como el anatomista alemán Gustav Schwalbe, a finales del siglo XX, existía un amplio consenso acerca de que Pithecanthropus erectus de Dubois era la especie ancestral que había dado lugar a Homo sapiens.
«La noción de que Homo sapiens podría haber compartido género con parientes extintos hoy en día no apareció hasta 1864»
Pero sorprendentemente pronto, y tal vez de manera inevitable, las interpretaciones comenzaron a cambiar. Al principio de la segunda década del siglo XX, el Pithecanthropus de Dubois se había vuelto una vez más claramente marginal, como se ve en la Figura 4, que muestra un diagrama filogenético humano publicado por el anatomista británico Sir Arthur Keith (1915). En esta concepción, casi todos los fósiles humanos o grupos de fósiles conocidos en aquel momento se veían como partes, o como el final, de linajes terminales. Resulta significativo que la mayoría de los que escribían sobre el registro fósil humano durante aquel período eran anatomistas humanos como Keith: científicos muy versados en los pequeños detalles de variación de Homo sapiens, pero con poco interés en los debates sobre mecanismos evolutivos entre genetistas, embriólogos, naturalistas y otros expertos abiertos durante las décadas posteriores al redescubrimiento en 1900 de los principios de la genética mendeliana. Los paleoantropólogos de este período parecen haber aceptado en general la noción darwinista de evolución como «descendencia con modificación», y dejan que otros se preocupen de los procesos subyacentes. Se limitaban a ocuparse de examinar los resultados. Sin embargo, a diferencia de la obligación a la que se enfrentaban los anatomistas comparativos, esto no implicó necesariamente tener que clasificar la diversidad de los seres vivos. Como resultado, y en lo que concernía a los paleoantropólogos, los nombres, incluso las denominaciones zoológicas formales, eran poco más que identificadores convenientes para fósiles individuales, más que una forma de perfilar unidades en la naturaleza. Sorprendentemente, esto se aplica tanto a los géneros como a las especies: para la mayoría de paleoantropólogos de este periodo, el nuestro era un género que contenía únicamente a Homo sapiens y a homínidos extintos con un tamaño cerebral similar, lo que a menudo incluía a los neandertales en el género Homo, pero no a muchas otras especies.
Para cuando llegó la Segunda Guerra Mundial, cualquier patrón discernible en un registro fósil que se expandía de forma constante había quedado oscurecido por una maraña de nombres informales y genéricos. Y sin importar la nomenclatura preferida, el árbol familiar homínido típico durante el periodo de entreguerras continuó, al estilo de Keith, representando la evolución homínida como un conjunto abundante de ramas terminales.
La síntesis evolutiva
El movimiento conocido como síntesis evolutiva nació durante las décadas de 1930 y 1940 a partir de una fusión teóricamente incómoda (aunque atractiva en términos reduccionistas) de la genética de poblaciones de base matemática y la teoría de la especiación. En última instancia, redujo la complejidad del proceso evolutivo a una simple cuestión de cambios graduales en las frecuencias con las que aparecían los genes en los linajes de organismos en evolución, guiados por la influencia de la selección natural. Especialmente en sus versiones más «duras», la síntesis mostraba que cada especie se transformaba gradualmente en su sucesora a lo largo de extensos períodos de tiempo. La aparición de los géneros y taxones superiores se veía simplemente como consecuencia pasiva de este tipo de transiciones repetidas.
«Para la mayoría, nuestro género contenía únicamente Homo sapiens y homínidos extintos con un tamaño cerebral similar»
Uno de los proselitistas más influyentes de la síntesis evolutiva fue el ornitólogo Ernst Mayr, quien fue más responsable que cualquier otro de acercar finalmente la teoría evolutiva a la paleoantropología. En 1950, Mayr publicó un artículo que reescribía literalmente las normas con las que debían operar los paleoantropólogos. Para entender la importancia de la revolución provocada por Mayr, cabe recordar que los relativamente escasos fósiles de homínidos conocidos en aquel momento se caracterizaban por una profusión de nombres de géneros y especies. De un plumazo, Mayr redujo unos quince nombres de géneros a uno solo. Es más, redujo literalmente docenas de nombres de especies a únicamente tres, y creó la siguiente clasificación de la familia homínida:
Familia Hominidae
Género Homo
Especie Homo transvaalensis [australopitecs]
Especie Homo erectus
Especie Homo sapiens [incluyendo los neandertales]
Finalmente, el género Homo volvía a estar en el centro de la paleoantropología. De hecho, la visión de Mayr abarcaba la familia Hominidae al completo. Es más, Mayr ubicó sus tres especies del género Homo en un único linaje evolutivo, volviendo a la linealidad pura del enfoque haeckeliano. Al proponer esta excesiva simplificación, Mayr tuvo muy poca consideración por el supuesto enfoque «no biológico» de los paleoantropólogos anteriores a la guerra, a los que directamente reprobó por no pensar en términos de variabilidad dentro de las poblaciones biológicas. Sorprendentemente, al menos en el entorno de habla inglesa, los paleoantropólogos aceptaron la contundente crítica de Mayr con una humildad asombrosa. De hecho, se precipitaron a unirse al carro del evolucionismo unilineal, posiblemente porque siempre habían sido conscientes subliminalmente del vacío teórico que ocupaba el centro de su disciplina. Tan profundos fueron los efectos del ataque de Mayr que, en menos de una década, nadie osaba utilizar nombres de géneros o especies en absoluto.
La preocupación de Mayr con la variación intraespecífica era, por supuesto, válida. Y se ajustaba cómodamente a la tradición anatómica humana de documentar la variación de Homo sapiens. Pero el nuevo enfoque también significaba que nadie se preocupaba en absoluto de las sinapomorfias morfológicas 3 que podrían haber mantenido al renacido género Homo como una unidad. En cambio, se crearon diagramas como el de la Figura 5, a partir de un artículo clásico escrito por el paleoantropólogo estadounidense Clark Howell (1957).
«La síntesis evolutiva nació a partir de la fusión de la genética de poblaciones de base matemática y la teoría de la especiación»
En el diagrama de Howell los participantes en el juego evolutivo son todos fósiles individuales, en su mayoría contenidos en una única masa filogenética sin forma que no evidencia de manera alguna la naturaleza de la diversidad entre los homínidos, o lo que podría unir al género Homo. Años después, los paleoantropólogos seguían publicando de forma rutinaria filogenias minimalistas de este tipo, con un tronco principal amplio, el de los homínidos, que daba lugar, como mucho, a un solo linaje secundario, aberrante y efímero: los robustos australopitecos, a quienes incluso Mayr (1953) se vio obligado a dejar de lado tras un exabrupto de John Robinson (1951). En estos constructos tan lineales, las especies –y por extensión los géneros– simplemente se clasificaban de manera inconsistente una tras otra a lo largo del tiempo. Y la consecuencia de ello era que resultaba imposible, casi por definición, reconocer especies y géneros en base a caracteres morfológicos derivados (incluso aunque alguien lo hubiera intentado). Los paleoantropólogos habían olvidado –si alguna vez lo comprendieron de verdad– el hecho de que los géneros, como agrupaciones de especies, son taxones superiores y que, en consecuencia, necesitamos definir el género Homo, como cualquier otro clado, basándonos en las sinapomorfias.
Después de la síntesis
En este contexto, no resulta sorprendente que la década de 1960 también fuera el período en el que se extendió la noción ultraminimalista de la evolución humana como «una única especie». Los antecedentes de esta noción son extraños y variados. Uno de ellos era Franz Weidenreich, el brillante anatomista y estudioso del hombre de Pekín (Weidenreich, 1939). Como la mayoría de sus contemporáneos, Weidenreich se vio poco afectado por cualquier teoría sobre cómo funcionaba realmente la evolución, aunque parece que se decantó por la ortogenesis, la noción de que la evolución está, de alguna manera, guiada por un objetivo. Esta idea era un anatema para los otros progenitores de la hipótesis de una sola especie, Theodosius Dobzhansky (1944) y Ernst Mayr (1950); no obstante, los tres miembros del trío se sentían cómodos con la idea de que la adquisición de la cultura había ampliado tanto el nicho ecológico del ser humano que, incluso por definición, no podría haber existido más de una especie humana en un momento dado del tiempo. Y si este fuera realmente el caso, significaría que, sin importar cuánto difieran entre ellos en términos anatómicos, todos los fósiles homínidos del mismo período tendrían que pertenecer necesariamente a la misma especie.
«El impresionante registro fósil del que disponemos hoy en día apunta realmente a una diversidad de especies en la historia de los homínidos»
Aunque a mitad de siglo ya era difícil basarse en morfologías fósiles, tal fue el impacto de Mayr para la paleoantropología que no fue hasta mitad de la década de 1970, cuando se demostró que dos tipos diferentes de homínido habían convivido en el mismo paisaje del norte de Kenia hace menos de dos millones de años, que los paleoantropólogos se vieron forzados una vez más a pensar en términos de diversidad sistemática entre los primeros homínidos. Desde ese momento, el registro fósil humano se acumuló rápidamente y evidenció que la historia de la familia humana no respondía solo al perfeccionamiento gradual de adaptaciones en un único linaje humano principal. Por el contrario, la historia de la familia homínida ha implicado constante experimentación evolutiva, con la aparición regular de nuevas especies que compiten en el escenario ecológico y en muchas ocasiones se extinguen. Y lejos de ser la cima de un árbol alto y esbelto, como consideraban Haeckel y Mayr, Homo sapiens es solo el final de una pequeña rama en un arbusto exuberante cuya complejidad se hace más y más evidente con el tiempo.
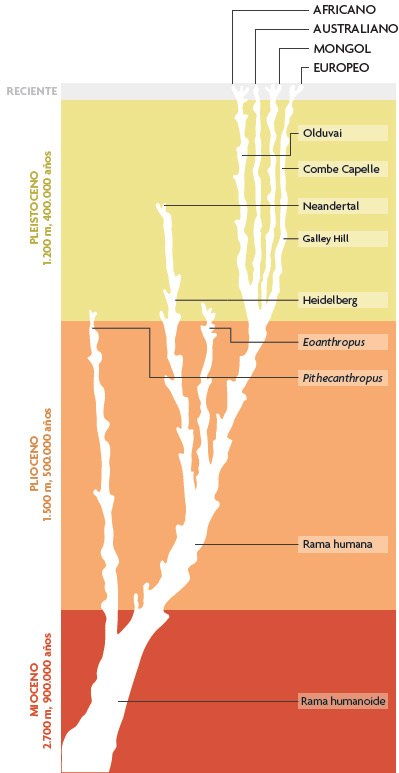
Figura 4. Filogenia de los homínidos propuesta por Sir Arthur Keith en 1915 en la que se señalan los fósiles humanos (o grupo de fósiles como los de los yacimientos de Combe Capelle o Olduvai) conocidos en aquel momento. / Redibujado por Kayla Younkin a partir de Theunissen, 1988
En 1993 publiqué una filogenia muy conjetural de la familia homínida (Tattersall, 1993), que muestra once especies de homínidos, y fue considerada bastante extravagante en aquel momento (Figura 6). Para cuando se redibujó el diagrama en 1998, la cuenta había ascendido a dieciséis. Curiosamente, muchas de las especies adicionales habían aparecido en la base del árbol, lo que demostraba que el patrón de diversidad y de experimentación evolutiva observado en el original había estado presente entre los homínidos desde el principio. En 2002, la imagen se había vuelto todavía más compleja, con veinte especies que había que encajar de alguna forma; y la versión más reciente (Figura 7) contiene 27 especies. Los detalles de estas filogenias tan inciertas, de igual manera que el número exacto de especies que se consideran, siguen siento tan discutibles como siempre. Pero lo que queda sobradamente claro es que no somos testigos únicamente de una explosión de nombres en un papel. El impresionante registro fósil de que disponemos hoy en día apunta realmente a una diversidad de especies en la historia de los homínidos.
El género ‘Homo’ hoy en día
Esta prueba de diversidad nos lleva de vuelta a las preguntas centrales que aborda este artículo. ¿Cómo se puede organizar correctamente la diversidad de los homínidos? ¿Qué parte de esta diversidad tiene cabida de forma legítima en nuestro propio género Homo? ¿Y cómo podemos reconocer los límites de este género? Como ya hemos señalado, un género es una agrupación de especies relacionadas por su ascendencia. Pero más allá del requisito general de ser monofilético, no existe un criterio general teórico ni práctico que dicte lo inclusivo que debería ser un género. Entonces, ¿dónde trazamos la línea?
A pesar de los esfuerzos de investigadores como Bernard Wood y Mark Collard (véase Collard y Wood, 2015; Wood y Collard, 1999), que se han pronunciado enérgicamente a favor de limitar el género Homo solo a homínidos que sean adaptativamente similares a la especie tipo (Homo sapiens) y que se relacionen más estrechamente a ella que al tipo de cualquier otro género, recientemente se tiende a la inclusión. De hecho, el ecuménico género Homo ya se ha extendido hasta hace 2,8 millones de años (Villmoare et al., 2015). Pero incluso suponiendo que la monofilia sea relevante en este caso, si queremos mantener cualquier valor como un repositorio de información filogenética o estructural, el género Homo resultante, amplio e inclusivo, requeriría un elaborado aparato de clasificación subgenérica. Lo que, por supuesto, da al traste con cualquier intento de agrupar a todos los homínidos relacionados. Por lo tanto, me parece mucho más práctico –además de más defendible en términos teóricos– comenzar con Homo sapiens e intentar conocer qué otras especies de homínidos sería útil clasificar en un género que está necesariamente definido por nuestra propia especie.
«Más allá del requisito general de ser monofilético, no hay un criterio general que dicte cómo de inclusivo debería ser un género»
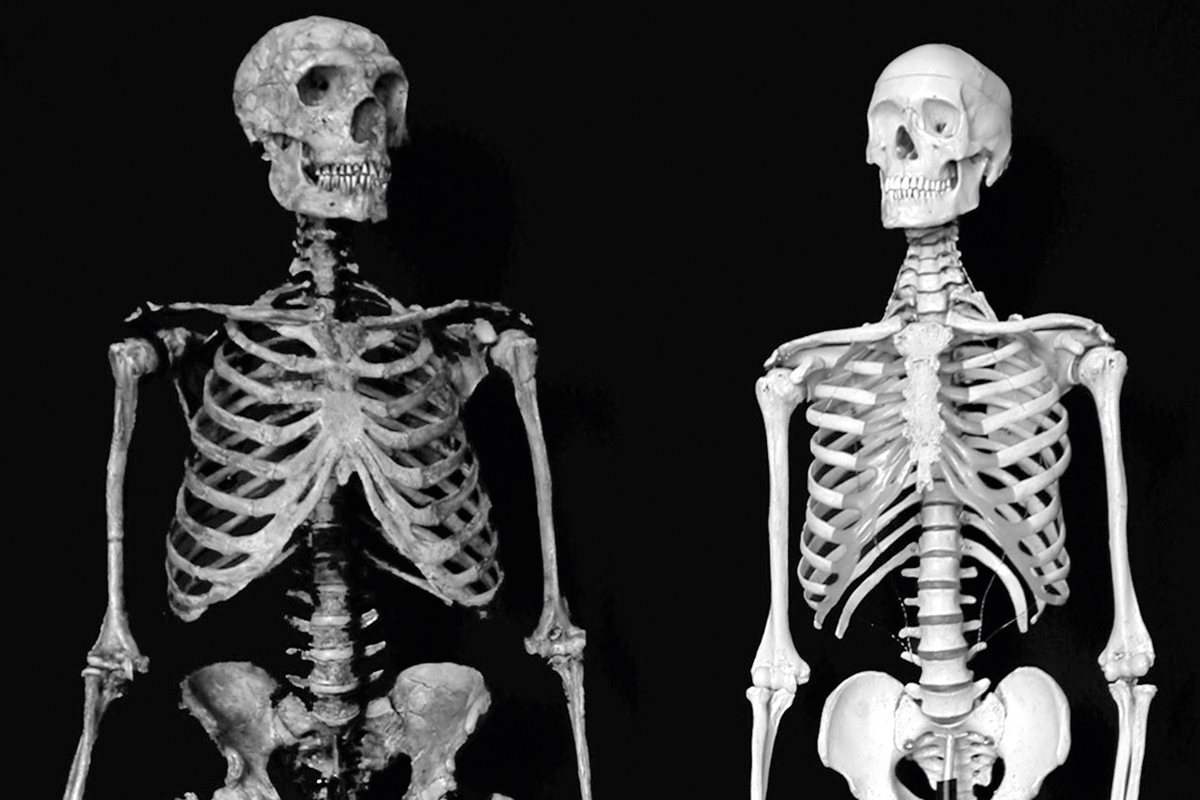
Inevitablemente, el número de especies que habría que reconocer entre los fósiles de homínidos más cercanos a Homo sapiens es un tema muy debatido. Por ejemplo, está claro que, dentro del grupo que se conocía a grandes rasgos como «Homo sapiens arcaico», queda representada una variedad de homínidos –aunque muy pocos considerarían hoy un problema incluir todas estas formas en el género Homo–. Lo mismo ocurre con los peculiares neandertales, hoy en día reconocidos de forma casi universal como Homo neanderthalensis. Estos homínidos europeos de gran cerebro están estrechamente relacionados con Homo sapiens, aunque ahora está claro que su clado se separó del linaje que lleva hasta Homo sapiens hace al menos medio millón de años (Arsuaga et al., 2014). También vale la pena mencionar en este contexto que la estructura osteológica de Homo sapiens parece haber derivado mucho: algo que resulta evidente en la comparación de la Figura 8 de un esqueleto de humano moderno con uno compuesto de neandertal que tiene un esqueleto postcraneal más primitivo. Una vez más, sin embargo, nadie discute actualmente que cualquiera de estas diferencias sean necesariamente un impedimento para la inclusión de ambos en el género Homo.
Si vamos más allá, el debate sigue abierto sobre qué debería constituir Homo erectus (la encarnación moderna del Pithecanthropus erectus de Dubois). Por un lado, se sostiene que esta especie es un taxón extendido por todo el viejo mundo, y que abarca una gran variedad de fósiles de homínidos de Asia y África a lo largo de casi dos millones de años. Otros, incluyéndome a mí, dirían que deberíamos considerar a Homo erectus una forma asiática endémica y terminal, en cuyo caso los homínidos africanos anteriores, a menudo asignados a Homo erectus –pero morfológicamente muy diferentes del tipo de Java–, deberían constituir su propia especie, Homo ergaster (Tattersall, 2015a). Pero otra vez parece no haber una razón teórica convincente por la que no podamos ver a todos los miembros del grupo erectus/ergaster como miembros de un género Homo filogenéticamente coherente. Porque, aunque algunos de ellos poseían cerebros que podían alcanzar un tamaño moderado (a lo sumo), todos parecen haber adquirido esqueletos de forma razonablemente moderna. De hecho, si los análisis y hallazgos futuros confirman que los homínidos están unidos por una configuración postcraneal común, quizás lo mejor sería reconocer el género Homo no solo mediante apomorfias craneales, sino también postcraneales. Esto podría resultar algo incómodo para los seres humanos modernos, que valoran lo que ocurre dentro de sus cabezas por encima de todo. Pero es posible que, al final, sea la única forma de reconocer un género Homo coherente en términos filogenéticos y estructurales.
Aun así, al incluir esta diversidad de fósiles ya estamos empezando a forzar los límites de lo que debería ser un género morfológicamente coherente; y si ampliamos el grupo erectus/ergaster para abarcar los sorprendentes hallazgos de homínidos de Dmanisi (Lordkipanidze et al., 2013), observamos un género Homo al que se ha forzado para acomodar una variedad poco convincente de tamaños cerebrales y morfologías craneales. Lo mismo ocurre incluso de forma aún más clara con «Homo» floresiensis de Indonesia –de una antigüedad inferior a los 100.000 años (Sutikna et al., 2016)– y con «Homo» naledi de Rising Star (Sudáfrica), que ahora creemos que data de hace entre 335.000 y 236.000 años (Dirks et al., 2017). Es más, incluso después de excluir del género Homo a los homínidos de Dmanisi, Flores y Rising Star, seguimos teniendo el conjunto de fósiles africanos de hace 1,6 millones de años o más, restos que en las últimas décadas se han atribuido a Homo habilis, a H. rudolfensis o simplemente a H. sapiens. Leakey et al. (2001) asignaron a rudolfensis a su nuevo género Kenyanthropus, mientras que Collard y Wood (2015) excluyeron (con buen juicio) todas estas formas de Homo. No obstante, un abrumador consenso sigue atribuyendo todos esos fósiles africanos antiguos a nuestro género.

Figura 5. Diagrama de Clark Howell de 1957 sobre la distribució temporal i espacial d’alguns homínids del plistocè mitjà i superior. Noteu que Howell no afirma en cap moment que això siga un «arbre». En blau, jaciments amb conjunts de fòssils humans. Redibuixat per Kayla Younkin / Adaptat per Salvador SastreFigura 6. Filogènia molt temptativa de la família Hominidae, proposada per Ian Tattersall en 1993. / Dibuixat per Diana Salles / Adaptat per Salvador Sastre
Cuando, profundamente influidos por la noción de «man the toolmaker» (“el hombre creador de herramientas”, véase Oakley, 1949), Louis Leakey y sus colegas llamaron Homo habilis a su nueva especie de hace 1,8 millones de años, proveniente de la garganta de Olduvai en Tanzania (Leakey, Tobias y Napier, 1964), lo hicieron siguiendo la tradición establecida de asignar formas fósiles a nuestro género basándose en comportamientos inferidos (dieta basada en carne, lenguaje, fabricación de herramientas, arte y simbolismo y demás), más que en la morfología observable. Sin embargo, su acción desató un gran furor en una ciencia que todavía era cautiva de Mayr. La mayoría de observadores o bien no querían separar estos fósiles de una especie de Homo erectus que lo abarcara todo, o bien rechazaban reconocer cualquier distinción significativa con respecto a los especímenes de Australopithecus de Sudáfrica. Los partidarios de esta segunda opinión tenían un argumento muy potente a su favor, puesto que parece evidente que la principal razón de Leakey y sus colegas para asignar los fragmentos de Olduvai a una nueva, aunque más «primitiva», especie de Homo era que estaban supuestamente asociados con las toscas herramientas de piedra encontradas en la capa más baja de la garganta.
Puesto que cualquier nuevo comportamiento, incluso algo tan rompedor como la fabricación de herramientas de piedra, tuvo que ser inventado necesariamente por un miembro de un género y una especie existente (de quien no deberíamos esperar diferencias biológicas significativas con respecto a sus padres o su descendencia), el criterio de Leakey y sus colegas para incluir a una especie en el género Homo nunca fue muy robusto. Sin embargo, por muchas razones (no muy convincentes), la noción de Homo habilis acabó cuajando. Además, un variado surtido de fósiles fragmentarios de hace, principalmente, entre 2 y 2,5 millones de años se atribuyeron al «primer Homo», especialmente después de que se descubrieran las primeras herramientas de piedra en un lugar que data de hace más de 2,5 millones de años (Semaw et al., 1997).
«La dinámica iniciada en Olduvai pronto dio lugar a una enérgica búsqueda del ‘primer Homo‘ que continúa aún hoy»
La dinámica iniciada en Olduvai pronto dio lugar a una enérgica búsqueda del «primer Homo» que continúa aún hoy. El último y más antiguo competidor por este puesto ha sido la forma Ledi-Geraru de Etiopía (Villmoare et al., 2015), con una edad de 2,8 millones de años. Sin embargo, ninguno de los pretendientes a «primer Homo» tiene muchas razones morfológicas para pertenecer a un género definido por el altamente derivativo Homo sapiens. En cambio, lo que parece ocurrir es que, aunque los paleoantropólogos no pueden evitar reconocer que existe diversidad en el registro de los primeros homínidos, siguen intentando dividir esa diversidad entre las dos categorías establecidas: Australopithecus y Homo. Y el deseo de embutir nuevas morfologías en esas dos casillas existentes cueste lo que cueste parece más fuerte que la evidente necesidad de ajustar el marco sistemático de los homínidos (con respecto a las especies y a los géneros) para dar cabida a nuevas morfologías.
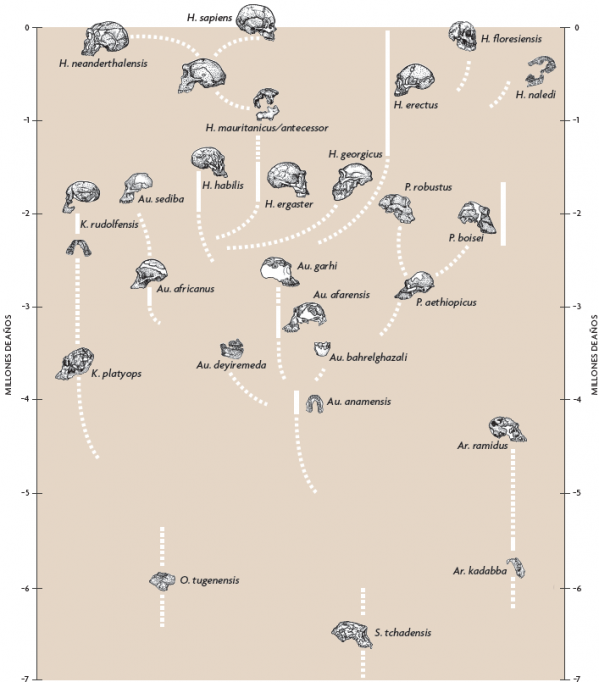
Figura 7. Filogenia muy tentativa de la familia Hominidae, propuesta por Ian Tattersall en 2017. Dibujado por Kayla Younkin / Adaptado por Salvador Sastre
Aun así, a pesar de la importancia de la tradición, ahora queda claro que en el pasado distante existieron múltiples linajes de homínidos. Y si queremos entender cómo se estructuraba esta diversidad, desde luego será mejor analizarlo en términos de ramificación de clados, en lugar de aferrarse de forma cada vez más aparatosa a la tradición de la división entre los dos géneros. Sí, en efecto, los géneros son clados; pero la diversidad morfológica que surge del creciente registro fósil homínido necesita claramente reflejarse en un contexto sistemático más extenso que el que permite la tradición (Schwartz y Tattersall, 2015; Tattersall 2015b, 2016). Al final, saturar un número limitado de géneros canónicos no ayudará a aclarar la evidentemente compleja serie de sucesos que llevó a la aparición de Homo sapiens.
Conclusión
A lo largo de la historia de la paleoantropología, el género Homo se ha contraído y expandido, no al servicio de la coherencia filogenética y morfológica, sino siguiendo patrones filosóficos. Hoy en día nos encontramos en un extremo de la expansión, con un concepto absolutamente ecuménico de género Homo que es, como mínimo, descuidado, tanto en términos filogenéticos como morfológicos. En un momento efímero hubo señales de que comenzaba una reevaluación tentativa del género y su contenido, con la transferencia de algunos especímenes icónicos de Homo primitivo a géneros como Australopithecus y Kenyanthropus (véase Leakey et al., 2001; Collard y Wood, 2015). Pero desde entonces el péndulo se ha atascado, por lo que todavía nos estorba un género que está literalmente abarrotado, tanto en el aspecto morfológico como en el sistemático.
«Ahora está claro que en el pasado distante existieron múltiples linajes de homínidos»
La lección que podemos aprender, o al menos deberíamos, es que el hecho de contener a Homo sapiens no convierte a Homo en un género especial. Por lo tanto, debemos abandonar nuestro excepcionalismo homínido y definir nuestro propio género exactamente igual que el resto. Lograr la coherencia de esta forma implicará reducir el género Homo, centrándose en reconocer un taxón firmemente basado por un lado en la sinapomorfia y que, por el otro, forma parte de un clado más complejo que necesita más géneros para expresar su diversidad. Esto, por supuesto, significa abandonar a aquellos candidatos extravagantes a «primer Homo» que solo lo son porque sus fósiles no son de australopitecinos.
1 Unidades morfológicas.(Volver al texto)
2 Parte más alta del cráneo que aloja el cerebro. (Volver al texto)
3 Una sinapomorfia es una novedad evolutiva, compartida por todos los individuos de un taxón que los diferencia a su vez de otros taxones.(Volver al texto)
REFERENCIAS
Arsuaga, J. L., Martínez, I., Arnold, L. J., Aranburu, A., Gracia-Téllez, A., Sharp, W. D., … Carbonell, E. (2014). Neandertal roots: Cranial and chronological evidence from Sima de los Huesos. Science, 344, 1358–1363. doi: 10.1126/science.1253958
Collard, M., & Wood, B. (2015). Defining the genus Homo. In W. Henke, & I. Tattersall (Eds.), Handbook of paleoanthropology (2n ed., vol. 3, pp. 2107– 2144). Heidelberg: Springer.
Dirks, P. H. G. M., Roberts, E. M., Hilbert-Wolff, H., Kramers, J. D., Hawks, J., Dosseto, A., … Berger, L. R. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife, 6e24231. doi: 10.7554/eLife.24231
Dobzhansky, T. (1944). On species and races of living and fossil man. American Journal of Physical Anthropology, 2, 251–265. doi: 10.1002/ ajpa.1330020303
Haeckel, E. (1868). Natürlische Schöpfungsgeschichte. Berlín: Reimer.
Howell, F. C. (1957). The evolutionary significance of variation and varieties of Neanderthal man. The Quarterly Review of Biology, 32, 330–347. doi: 10.1086/401978
Keith, A. (1915). The antiquity of man. Londres: Williams and Norgate.
King, W. (1864). The reputed fossil man of the Neanderthal. Quarterly Journal of Science, 1, 88–97.
Leakey, L. S. B., Tobias, P. V., & Napier, J. R. (1964). A new species of Homo from Olduvai Gorge. Nature, 202, 7–9. doi: 10.1038/202007a0
Leakey, M. G., Spoor, F., Brown, F. H., Gathogo, P. N., Leakey, L. N., & McDougall, I. (2001). New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature, 410, 433–440. doi: 10.1038/35068500
Linnaeus, C. (1758). Systema Naturae (10ª ed.). Estocolmo: Salvii.
Lordkipanidze, D., Ponce de León, M. S., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A., & Zollikofer, C. P. E. (2013). A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 342, 326–331. doi: 10.1126/science.1238484
Mayr, E. (1950). Taxonomic categories in fossil hominids. Cold Spring Harbor Symposia on Quantitative Biology, 15, 109–118. doi: 10.1101/ SQB.1950.015.01.013
Mayr, E. (1953). Comments on evolutionary literature. Evolution, 7(3), 273– 281. doi: 10.2307/2405739
Oakley, K. P. (1949). Man the toolmaker. Londres: Trustees of the British Museum.
Robinson, J. T. (1951). Meganthropus, australopithecines and hominids. American Journal of Physical Anthropology, 11, 1–38. doi: 10.1002/ ajpa.1330110112
Schwartz, J. H., & Tattersall, I. (2015). Defining the genus Homo. Science, 349, 931–932. doi: 10.1126/science.aac6182
Semaw, S., Renne, P., Harris, J. W. K., Feibel, C. S., Bernor, L., Fesseha, N., & Mowbray, K. (1997). 2.5 million-year-old stone tools from Gona, Ethiopia. Nature, 385, 333–336. doi: 10.1038/385333a0
Sutikna, T., Tocheri, M. W., Morwood, M. J., Saptomo, E., Jatmiko, Awe, R. D., … Roberts, R. G. (2016). Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature, 532, 366–369. doi: 10.1038/nature17179
Tattersall, I. (1993). The human odyssey. Englewood Cliffs, NJ: Prentice-Hall.
Tattersall, I. (2015a). Homo ergaster and its contemporaries. In W. Henke, & I. Tattersall (Eds.), Handbook of paleoanthropology (2ª ed., vol. 3, pp. 2167–2188). Heidelberg: Springer.
Tattersall, I. (2015b). Defining and recognizing the genus Homo. Gortania, 36, 5–22.
Tattersall, I. (2016). The genus Homo. Inference: International Review of Science, 2(1). Consultado en http://inference-review.com/article/the-genus-homo
Theunissen, B. (1988). Eugéne Dubois and the Ape-Man from Java: The history of the first “missing link” and its discoverer. Dordrecht/Boston: Kluwer Academic.
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, E. N., DiMaggio, E. N., Rowan, J., … Reed, K. E. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science, 347, 1352–1355. doi: 10.1126/science. aaa1343
Weidenreich, F. (1939). Six lectures on Sinanthropus pekinensis and related problems. Journal of the Geologicual Society of China, 19, 1–110. doi: 10.1111/j.1755-6724.1939.mp19001008.x
Wood, B., & Collard, M. (1999). The human genus. Science, 284, 65–71. doi: 10.1126/science.284.5411.65




