

|
The intelligent design delusion. Obsolete science and pseudoscience in neocreationism. After the 1987 sentence of the US Supreme Court against teaching creationism in the biology classroom, the intelligent design movement sentenced Darwin’s failure in the biochemical realm. At any rate, this new assault on scientific scrutiny of the living world is none other than the old pseudoscience of creationism wrapped up in inaccurate biochemistry «La ignorancia engendra atrevimiento más frecuentemente que la ciencia: son los que saben poco, y no los que saben mucho, los que afirman positivamente que esto o lo otro no lo podrá resolver nunca la ciencia». Charles Darwin (1871) Los factores culturales fueron determinantes en la manera cómo los científicos de diferentes países sintonizaron o no con la aproximación de Charles Darwin al origen de la complejidad y diversidad biológica. Hace poco más de un siglo, para algunos científicos católicos el dilema definitivo sobre el origen de la vida era o la generación espontánea –un postulado acientífico tras los experimentos contundentes de Louis Pasteur y John Tyndall– o la Creación. Para ellos –notablemente el entomólogo alemán y jesuita Erich Wasmann, polemista de Ernst Haeckel–, esta aproximación neovitalista era totalmente compatible con un proceso evolutivo de los seres vivos. A pesar de todo, la Iglesia católica, lastrada por el caso Galileo, evitó la confrontación directa y la condena pública y oficial del evolucionismo. Se limitó en un primer momento a amonestar uno a uno los pocos autores católicos favorables a la evolución, sin hacer mucho ruido. El caso de algunas confesiones protestantes es muy diferente. Si bien los grupos más liberales aceptaron a Darwin pronto, durante el siglo pasado los grupos cristianos fundamentalistas norteamericanos, defensores de la literalidad bíblica, han tenido como objetivo prioritario la introducción de la narración del Génesis en la clase de biología, como alternativa a la teoría evolucionista. El fracaso del asalto antinaturalista a la educación científica se produjo a finales de la década de 1980, dados los motivos religiosos demasiado evidentes del llamado «creacionismo científico» y la nítida separación Iglesia-Estado consagrada por la constitución norteamericana. Pese a ello, la sentencia de 1987 del Tribunal Supremo de EEUU contra el creacionismo fue el catalizador de la emergencia en los años noventa de una forma más sutil conocida como «diseño inteligente» (DI). Algunos la han llamado el caballo de Troya del creacionismo de siempre y el juez John Jones, que sentenció su carácter religioso y no científico a finales de 2005, calificó la decisión del consejo escolar de Dover (Pensilvania) de imponer la enseñanza de esta teoría en la clase de ciencias como una «inanidad imponente» y una vergonzosa pérdida de tiempo para el sistema judicial. La propuesta del DI se basa en conceptos como el de la «complejidad irreducible» de las estructuras biomacromoleculares. El bioquímico de la Universidad de Lehigh Michael Behe presenta esta noción neocreacionista como una alternativa a la evolución por selección natural. Pero así como Wasmann y sus seguidores tenían como punto de referencia la excelencia científica de Pasteur, Behe sólo se basa en la ignorancia, la supuesta incapacidad actual de explicar el origen y evolución de determinadas estructuras celulares. Conviene señalar que no hay nada nuevo en la fascinación por el diseño aparente en la naturaleza porque, aunque no lo reconozcan, los defensores del DI son descendientes intelectuales del reverendo William Paley. Simplemente, trasladan la admiración ante la estructura y adaptación de un ojo a las intimidades subcelulares y eligen como súmmum de la complejidad la motilidad bacteriana. Si el proceso contra el maestro rural que explicaba evolución, inmortalizado en la película Inherit the wind, fue conocido como el «juicio del mono», el de Dover ha sido el «juicio del flagelo bacteriano». |
«La Iglesia Católica, lastrada por el caso galileo, evitó la confrontación directa y la condena pública y oficial del evolucionismo» | |
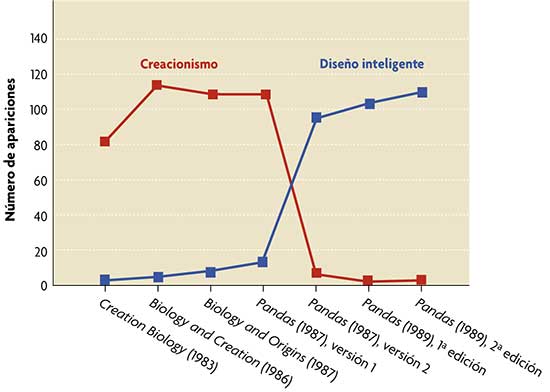
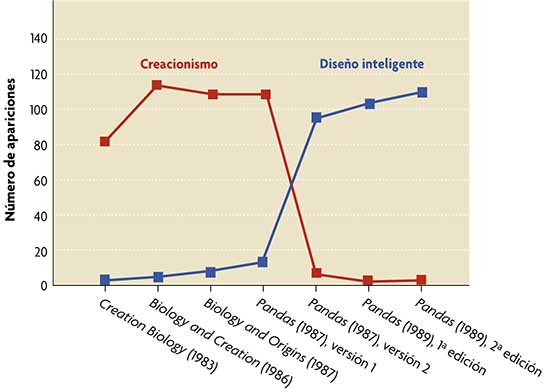 El término «diseño inteligente» desplazó las referencias directas al creacionismo en el principal libro divulgativo de este movimiento, Of pandas and people. Eso ocurrió tras la sentencia de 1987 del Tribunal Supremo de los EEUU contra la enseñanza del creacionismo en la escuela pública. Los manuscritos iniciales usaban profusamente términos derivados de creación (gráfico en rojo) que sistemáticamente fueron sustituidos por diseño inteligente en ediciones posteriores del libro (gráfico en azul). Para comparar, se incluyen textos creacionistas anteriores a 1987. La historiadora de la ciencia Barbara Forrest desenmascaró esta burda maniobra durante el juicio de Dover, lo que constituyó una prueba definitiva para el juez Jones de que «el DI ha evolucionado a partir de formas ancestrales de creacionismo.» El gráfico es una adaptación, con permiso de los autores, de la figura 1 de: Scott, E. C. y N. J. Matzke, 2007. «Biological design in science classrooms». Proc. Natl. Acad. Sci. USA, 104: 8669-8676. |
«En el siglo pasado, el objetivo prioritario de los grupos cristianos fundamentalistas norteamericanos fue la introducción del Génesis en la clase de biología» | |
|
Sin embargo, un análisis bioquímico detenido de la noción de complejidad irreducible y de otras afirmaciones extravagantes de los defensores del DI nos muestra que ésta –como las otras aproximaciones vitalistas a los fenómenos biológicos– no es una hipótesis testable científicamente. Más bien nos encontramos ante una bioquímica obsoleta e incorrecta que sirve de disfraz a la seudociencia del creacionismo. El argumento de la complejidad irreducible Darwin’s black box: the biochemical challenge to evolution es un libro de Behe publicado en 1996 (Free Press, Nueva York) y a menudo se considera una de las obras centrales del neocreacionismo del DI. Introduce el concepto de complejidad irreducible: esta propiedad, afirma Behe, la tienen aquellas estructuras biológicas complejas, formadas por muchas piezas interdependientes, de manera que la función del conjunto se pierde con sólo que eliminemos una pieza. Para Behe estas estructuras no pueden haber emergido por un proceso de selección natural, porque encuentra inimaginable la existencia de estadios intermedios, con menos piezas, que carecerían de función. Su conclusión es que estas complejidades han sido diseñadas por una entidad no natural, sin explicitar de qué tipo. El concepto de complejidad irreducible depende en buena medida de lo incompleto de nuestro conocimiento de los mecanismos evolutivos, una ignorancia lógicamente pasajera. Por lo tanto, y a diferencia de la idea del DI, la complejidad irreducible de una estructura celular es refutable. Veamos el caso del icono por excelencia del DI: el flagelo bacteriano, el aparato de locomoción de muchos microorganismos. Para empezar, tenemos que referirnos, como hace Behe, a que «el flagelo» bacteriano esconde una concepción esencialista de la biología celular que no se corresponde con la realidad. Hay una diversidad abrumadora de aparatos de motilidad entre los microorganismos y tenemos un conocimiento bioquímico pormenorizado de unos pocos ejemplos. La genómica ambiental revela que debe haber millones de sistemas de motilidad diferentes en el mundo microbiano. Dado que la ciencia microbiológica afecta a una fracción insignificante de la biodiversidad microscópica de la naturaleza, el argumento de Behe se basa en un conocimiento provisional y fragmentario de la motilidad bacteriana. Éste es un frágil fundamento para sostener un argumento tan fuerte como que la alternativa explicativa para la evolución natural del flagelo es la DI. La consecuencia inmediata es que el Dios obsesionado con los escarabajos de John B. S. Haldane se convierte en una divinidad dedicada a diseñar motores moleculares. Nos percatamos así hasta qué punto los partidarios del DI subvierten el procedimiento científico. Una hipótesis científica se debe contrastar con hechos y observaciones que la apoyen o la rechazen pero nunca se debe basar en la incapacidad explicativa de otra hipótesis. En el caso del esquema evolutivo, la explicación es incompleta porque nuestro conocimiento de la naturaleza y sus mecanismos es incompleto y eso no implica en absoluto que no llegaremos a mejorar nunca estos esquemas explicativos. Sin embargo, para desgracia de Behe y de sus partidarios, en el caso del flagelo sí que tenemos un esquema evolutivo que, aun siendo provisional, es satisfactorio. Irónicamente, es el mismo que ya sugirió el mismo Darwin hace ciento cincuenta años para abordar la evolución del ojo de los animales. La selección natural ha actuado sobre estructuras más simples que eran adaptativas para los seres que las poseían, es decir, que eran heredables y aumentaban las posibilidades de supervivencia y reproducción de éstos ante otros que carecían de ellas. Si bien durante la evolución de los ojos la presión selectiva ha sido la capacidad de detectar la luz solar y de captar imágenes, los antepasados de los flagelos bacterianos no tenían una función motora: eran maquinarias de secreción proteica. De hecho, determinados sistemas de secreción actuales son homólogos (es decir, comparten antepasados evolutivos comunes) con componentes fundamentales de la maquinaria motora. He aquí una de las dificultades que afronta la reconstrucción del pasado: los predecesores de una determinada estructura quizá no tenían la misma función que sus descendientes. Además, la evolución por selección natural, como propuso François Jacob, extendiendo a escala molecular una noción del mismo Darwin, se parece más a la manera de proceder del bricolaje que al plan de un ingeniero. La evolución es oportunista y combina partes con unas funciones originales que no coinciden necesariamente con la función que emerge del conjunto final. Por precaria que sea la nueva función, si proporciona una ventaja reproductiva, continuará existiendo y pasando de generación en generación, eventualmente sometida a procesos de refinamiento y optimización. Desde el punto de vista de la bioquímica, el argumento de la complejidad irreducible contiene otro error colosal. Behe considera que las proteínas tienen una única función y, por lo tanto, como componentes de edificios macromoleculares, tienen una determinada funcionalidad relacionada con el conjunto. Esta visión esencialista y platónica de las proteínas es falsa. Las proteínas son polifacéticas. Una proteína que forma parte de un complejo puede ejercer otra función por ella misma, aisladamente o en conjunción con otras. Esta realidad es ignorada por Behe y esto invalida de arriba a abajo el argumento de la complejidad irreducible, porque los componentes aislados sí que pueden ser funcionales y representar una ventaja adaptativa para su poseedor. El problema actual es que sabemos muy poco de las diversas ocupaciones de las proteínas. Es una realidad que ha empezado a emerger y que crece ante nosotros cada día. No podemos, pues, renunciar a la investigación científica de estas múltiples funciones y al análisis comparado en el mayor número posible de organismos para mejorar nuestros esquemas evolutivos. La única prueba a favor del DI que nos aporta Behe es su vagancia intelectual. La renuncia al naturalismo metodológico Si bien ha quedado claro que los autores del DI como alternativa de la teoría evolutiva hacen trampa o, por lo menos, exhiben desvergonzadamente la debilidad de sus conocimientos bioquímicos, hay también una cuestión de fondo. Por lo que respecta a la identidad del diseñador, es cierto que los proponentes del DI intentan no hablar de Dios –del Dios personal, omnipotente y omnisciente de la tradición judeocristiana, claro. Incluso se ha llegado a decir que el diseñador podría ser un biólogo molecular viajero del tiempo. Pero en ciertas ocasiones, como durante el juicio de Dover, se ha reconocido explícitamente que la plausibilidad del argumento del DI depende de hasta qué punto uno crea en la existencia de Dios. Y ello se convirtió en una prueba irrefutable del carácter religioso y no científico de la propuesta, como señaló el juez Jones. Así, dado el carácter sobrenatural del diseñador, la venta y difusión publicitaria del DI como una alternativa científica de la teoría evolucionista requiere una redefinición de las fronteras de la ciencia. Estos autores piden que la ciencia incluya otras causas que no sean naturales. Eso significa una renuncia radical y absoluta a lo que ha sido el método científico en los últimos siglos. Independientemente de las creencias o las ideologías personales, los científicos siempre han explorado las causas naturales de los fenómenos sometidos a estudio. Este naturalismo metodológico es el fundamento de todo nuestro conocimiento científico de la naturaleza y de sus aplicaciones tecnológicas. El ataque al propio concepto de ciencia también supondría transmutar en disciplina científica, como aseguran los neocreacionistas, materias como la astrología. La obsesión por mezclar la religión y la ciencia desemboca en estas extravagancias. Sin embargo, como ya propuso el evolucionista teísta Theodosius Dobzhansky, y hace poco ha reiterado Ayala, el creacionismo es una blasfemia. Según algunos teólogos, la teoría de la evolución es un regalo porque permite liberar al Creador de la carga de tanta chapucería y crueldad como abunda en la naturaleza. El DI no se puede separar de sus antepasados religiosos y creacionistas y, por lo tanto, se trata de una nueva incursión del vitalismo, la magia y la mitología, esta vez con un disfraz tejido con una bioquímica de muy baja calidad. Por suerte, esta vestimenta es fácil de arrancar y no puede esconder la vergüenza de una visión oscurantista de la naturaleza con la que no tenemos absolutamente nada que ganar pero sí muchísimo que perder. BIBLIOGRAFÍA PARA PROFUNDIZAR Juli Peretó. Departamento de Bioquímica y Biología Molecular e Institut Cavanilles de Biodiversitat i Biologia Evolutiva, Universitat de València. |
«Una hipótesis científica se debe contrastar con hechos y observaciones, pero nunca se debe basar en la incapacidad explicativa
«Los proponentes del diseño inteligente intentan no hablar de Dios. Se ha llegado a decir incluso que el diseñador podría ser un biólogo molecular viajero del tiempo»
«El creacionismo es una blasfemia. La teoría de la evolución permite liberar al Creador de la carga de tanta crueldad como abunda en la naturaleza» |
 © Nicolle Rager Fuller, National Science Foundation Escherichia coli en movimiento. |
||
| De secretar a navegar
El flagelo representa el principal mecanismo de la motilidad bacteriana. Tras este nombre, sin embargo, se esconde una extraordinaria diversidad de sistemas de motilidad en microorganismos. El arquetipo del flagelo es el de Salmonella typhimurium y Escherichia coli, una estructura ciertamente compleja que depende de la actividad de 30 genes. Los principales constituyentes son un propulsor o filamento –una larguísima sucesión de más de 20.000 unidades de la proteína llamada flagelina–, conectado a través de un gancho al cuerpo basal, formado por unas 500 copias de varias proteínas. Las diversas partes tienen funciones bien definidas: el complejo de exportación (incluyendo el ATPasa, una enzima que facilita energía para la salida de la flagelina constituyente del filamento), varios anillos de fijación a las membranas y la pared celulares, y el motor que propulsa el filamento. El cuerpo basal también dispone de un cambio de marchas que al variar el sentido de la rotación del motor permite a la bacteria cambiar de dirección. A su vez el motor consta de un estátor –una estructura que permanece estática sumergida en la membrana interna y anclada en la pared– y de un rotor (también llamado anillo C). El combustible de la motilidad es un flujo de iones de hidrógeno (protones) que pasa a través del rotor, del exterior hacia el interior celular, y le imprime un movimiento de rotación que se transmite al filamento por medio de la biela. |
||
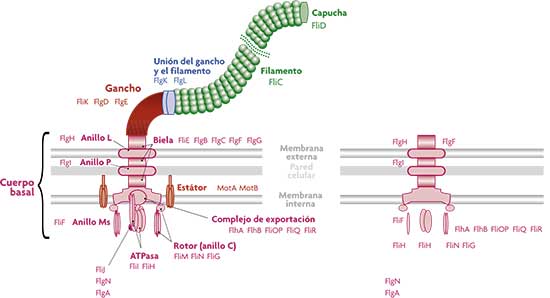 Los esquemas (a la izquierda, esquema del flagelo E. Coli y, a la derecha, esquema de los vestigios del flagelo en Buchnera aphidicola) son adaptaciones de la figura 20 de: «Secuenciación y análisis del genoma completo de Buchnera aphidicola BCc, endosimbionte primario del pulgón Cinara cedri (Aphididae: Lachninae)», Vicente Pérez-Brocal, tesis doctoral, Universitat de València, 2006. Los esquemas (a la izquierda, esquema del flagelo E. Coli y, a la derecha, esquema de los vestigios del flagelo en Buchnera aphidicola) son adaptaciones de la figura 20 de: «Secuenciación y análisis del genoma completo de Buchnera aphidicola BCc, endosimbionte primario del pulgón Cinara cedri (Aphididae: Lachninae)», Vicente Pérez-Brocal, tesis doctoral, Universitat de València, 2006. |
«En las bacterias Buchnera observamos un vestigio del pasado que el mismo Darwin consideraría como prueba de la evolución» | |
|
En las bacterias endosimbiontes de insectos, como Buchnera aphidicola, que habita en ciertas células especializadas de los pulgones, se observa una extraordinaria reducción genómica durante la adaptación a la vida intracelular. Buchnera aphidicola tiene un antepasado común con E. coli. La secuenciación completa del genoma del endosimbionte primario del pulgón del cedro Cinara cedri muestra que en los últimos 250 millones de años la bacteria ha perdido muchos de los componentes del flagelo de sus antepasados. Las estructuras remanentes constituyen una maquinaria similar al sistema de secreción de proteínas de tipo III, un dispositivo que se considera como el precursor evolutivo del flagelo. Por lo tanto, en Buchnera observamos un atavismo, un vestigio del pasado que el mismo Darwin consideraría como prueba de evolución: la pérdida de la motilidad, obviamente asociada a la vida intracelular, ha dejado atrás una porción del flagelo especializada en la secreción de proteínas. Esta es una demostración directa de que el flagelo no representa un caso de «complejidad irreducible» como sostiene Behe, o de «relación» entre partes, que diría Paley. Bien mirado, en el flagelo actual la secreción de proteínas es una función esencial durante su propia biogénesis: el propulsor del aparato está constituido por proteína secretada en el exterior que espontáneamente polimeriza y forma el filamento. MAS INFORMACIÓN Juli Peretó. Departamento de Bioquímica y Biología Molecular e Institut Cavanilles de Biodiversitat i Biologia Evolutiva, Universitat de València. |
||




