

La evolución del género ‘Homo’
Nuevas incógnitas y perspectivas

En este trabajo se pasa revista a las principales cuestiones que afectan a la evolución del género Homo, como es el caso de su origen, el problema de la variabilidad en Homo erectus y el impacto de la paleogenómica. Todavía no se ha llegado a un consenso sobre el candidato australopitecino que dio lugar a los primeros representantes asignables a Homo. Esta discusión afecta incluso al propio reconocimiento de las especies H. habilis y H. rudolfensis. Por lo que respecta a la cuestión de la variabilidad de los primeros paleodemes asignados a Homo, los hallazgos realizados en el yacimiento georgiano de Dmanisi han puesto en cuestión algunos de los criterios utilizados hasta ahora para distinguir entre especies como H. erectus o H. ergaster. Finalmente, la irrupción de la paleogenómica ha puesto en evidencia que el flujo genético entre antiguas poblaciones de homininos fue mayor de lo esperado.
Palabras clave: paleogenómica, género Homo, homininos, variabilidad, Dmanisi.
En los últimos años, nuestra concepción del origen y evolución de nuestro género se ha visto sacudida por diferentes hallazgos que, lejos de dar respuesta a los problemas que se planteaban a finales del siglo XX, han reabierto debates y han obligado a replantearse los esquemas que se mantuvieron vigentes durante décadas. Algunas de estas cuestiones abiertas lo son porque faltan los fósiles adecuados que nos den la respuesta. Otras, por el contrario, son todavía objeto de debate por la diferente interpretación que se da a algunos fósiles y a la adscripción sistemática de estos. Hallazgos como los del yacimiento georgiano de Dmanisi o los proporcionados por la paleogenómica ponen en cuestión, sobre todo, los criterios utilizados hasta ahora para reconocer especies en el registro fósil. A continuación enumeramos algunas de estas cuestiones.
El origen del género ‘Homo’
Los primeros representantes de nuestro género hacen su aparición en el registro fósil africano hace unos 2,8 millones de años (Villmoare et al., 2015). Estos primeros restos de nuestro género han sido atribuidos en ocasiones a Homo rudolfensis. Tenemos evidencias de una segunda especie, Homo habilis, de hace algo más de dos millones de años, que se diferencia de la anterior en algunos caracteres secundarios y en la menor capacidad craneana, aunque para algunos investigadores Homo habilis y Homo rudolfensis corresponden en realidad a la misma especie.
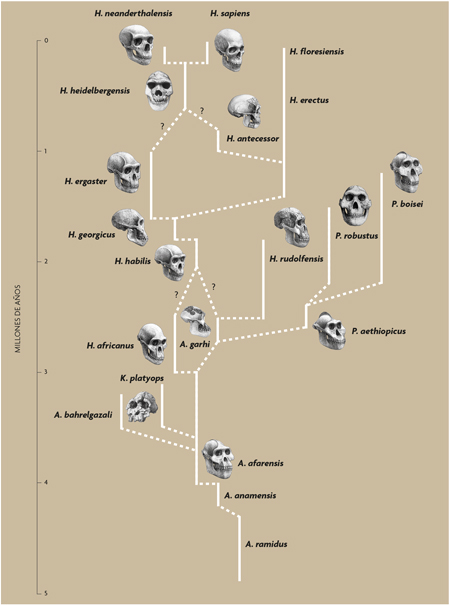
Esquema de la evolución humana propuesta por Agustí y Lordkipanidze (2005). En los últimos años, nuestra concepción del origen y evolución del género Homo se ha visto sacudida por diferentes hallazgos que han reabierto debates y obligado a replantearse ciertos esquemas. / Ilustraciones de los cráneos: Mauricio Antón
Hasta mediados de los años setenta del pasado siglo, existía un claro candidato australopitecino para ocupar el puesto de ancestro de nuestro género, Australopithecus africanus, de distribución exclusivamente sudafricana. La publicación en 1978 de los restos de Australopithecus afarensis procedentes de Afar, en Etiopía, supuso el primer cuestionamiento de esta filiación. Para sus descubridores, Don Johanson y Tim White, Australopithecus afarensis, con una cronología de entre 3,5 y 3 millones de años, encajaba como perfecto antecesor de nuestro género, mientras que Australopithecus africanus habría dado lugar en Sudáfrica a las formas robustas de australopitecinos encuadradas en el género Paranthropus.
Con posterioridad, en 1996, el mismo White propuso una nueva especie, Australopithecus garhi, encontrada en niveles datados en unos 2,5 millones de años en la localidad de Bouri, en Etiopía. Según White y sus colaboradores, A. garhi se encontraba en el lugar y en el momento adecuado para ser el ancestro de nuestro género. A ello se añadió el hecho de haber encontrado asociados a A. garhi huesos que presentaban marcas de corte de útiles líticos, de los cuales se supone que esta especie fue la responsable.
Para acabar de complicar el tema, en 2001 el equipo liderado por Meave Leakey propuso un nuevo género, Kenyanthropus, para un cráneo muy deformado procedente de la cuenca de Lomekwi en Kenia, y datado en unos 3,5 millones de años (esto es, contemporáneo de A. afarensis). La capacidad craneal estimada del llamado Kenyanthropus platyops era baja, en torno a 350 cm3. Pero lo que llevó a sus descubridores a proponer un nuevo género fue la cara, larga y plana. Dando un paso más allá, plantearon que esta forma era el antepasado de Homo rudolfensis, especie que propusieron incluir también en el género Kenyanthropus. Esta última proposición ha hallado escaso eco entre la comunidad paleoantropológica, que, lejos de aceptar el nuevo género, ha optado por incluir la especie K. platyops dentro de Homo o de Australopithecus. Sin embargo, algunos autores sí que han aceptado la sugerencia de una relación entre K. platyops y H. rudolfensis.
Más allá de los hallazgos en el este de África, la idea de un origen sudafricano de nuestro género volvió a saltar a la palestra con el descubrimiento de una nueva especie de Australopithecus, A. sediba, en la cueva de Malapa en Sudáfrica. Se trata de dos esqueletos parciales pertenecientes a una mujer joven de unos veinte años y a un adolescente de unos doce. El elemento más significativo es un cráneo en excelente estado de conservación que revela un Australopithecus próximo a Australopithecus africanus pero con caracteres más derivados. Aparte de que poseía un cerebro algo mayor que el de su predecesor, la cara muestra algunos rasgos que lo aproximan a los primeros representantes de nuestro género, como Homo habilis (por ejemplo, la forma de la cavidad nasal y los pómulos). Para su descubridor, Lee Berger, de la Universidad de Witwatersrand (Sudáfrica), Australopithecus sediba constituye el perfecto eslabón intermedio entre Australopithecus africanus y los primeros representantes del género Homo. El problema, sin embargo, reside en que los restos de Australopithecus sediba están fechados en unos 1,8 millones de años y son, por tanto, muy posteriores a la eclosión de nuestro género, que data de más de 2,5 millones de años. Se debe a este mismo equipo el reconocimiento de una nueva especie arcaica del género Homo, H. naledi, en Dinaledi Chamber, una cavidad de muy difícil acceso también en Sudáfrica. Sin embargo, la ausencia de un adecuado contexto geológico y paleontológico cuestiona la posible relevancia de este hallazgo en relación con el origen de nuestro género.

Homo rudolfensis ha sido considerado como el primer representante del género Homo, cuyos restos se remontarían a hace unos 2,8 millones de años. Hace poco más de dos millones de años, tendríamos evidencias de la existencia de una segunda especie, Homo habilis, aunque algunos investigadores consideran que tanto H. rudolfensis como H. habilis son la misma especie. En la imagen, cráneo y reconstrucción de Homo rudolfensis (según Mauricio Antón). / Ilustración: Mauricio Antón
En relación a los candidatos australopitecinos del este de África, existe un dato sorprendente y es la tendencia a poseer grandes molares (un fenómeno conocido como «megadoncia»), que llegará a su máximo exponente con las especies incluidas en el género Paranthropus. Esta tendencia es ya evidente en Australopithecus afarensis, especie que, si bien podría cuadrar como un buen antepasado de Homo, igualmente encajaría como un plausible ancestro de Paranthropus. De hecho, cuando salió a la luz el llamado «cráneo negro» de Paranthropus aethiopicus, diversos autores hicieron notar sus afinidades con Australopithecus afarensis. Si vamos ahora a Australopithecus garhi, esta tendencia a la megadoncia se hace todavía más patente, aunque la morfología craneal no permita incluirla en Paranthropus. Sin embargo, es evidente que en el este de África estamos en un contexto ambiental que favorecía el desarrollo de un potente aparato masticador y grandes molares.
Cuando a continuación pasamos a analizar la dentición de los primeros miembros de nuestro género, nos encontramos con la sorprendente conclusión de que también los primeros restos asignados a Homo poseían grandes molares, razón por la cual son comúnmente incluidos dentro de la especie Homo rudolfensis. Solo más tardíamente aparecen los restos asignados a Homo habilis, cuya dentición era más grácil y próxima a la de los Australopithecus sudafricanos.
Estamos, por tanto, ante una dicotomía. Así, en África oriental podríamos reconocer un linaje que, a través de Australopithecus afarensis y Australopithecus ghari (y tal vez Kenyanthropus), conduce a Homo rudolfensis. Por otro lado, existiría una línea grácil que desde Australopithecus africanus llevaría a Homo habilis, tal vez incluyendo a Australopithecus sediba. La sorprendente conclusión sería un origen difilético para el género Homo.
Ahora bien, ante un registro tan fragmentario como el correspondiente a las primeras variantes del género Homo, cabe preguntarse si en realidad tanto Homo habilis como Homo rudolfensis corresponden a variantes dentro de una misma especie. Los elementos diferenciales entre ambas pueden ser una consecuencia de la propia variabilidad poblacional dentro de esa especie.
El estatus de ‘Homo erectus’ y el tema de la variabilidad
A mediados de los años setenta del siglo pasado, dos revoluciones conceptuales en el campo de la biología evolutiva iban a influir profundamente en las ideas sobre nuestra evolución. De una parte, en la sistemática paleontológica se extendió el recurso a la cladística a la hora de establecer la filogenia de cualquier grupo fósil. De esta manera, con su distinción entre caracteres primitivos y derivados, se pudieron elaborar hipótesis contrastables sobre el parentesco entre diferentes taxones.

En 2001, el equipo de la paleoantropóloga Meave Leakey propuso el nuevo género Kenyanthropus. Para estos investigadores, Kenyanthropus platyops era el antepasado de Homo rudolfensis, propuesta que ha contado con escaso eco en la comunidad científica. En la imagen, cráneo y reconstrucción de Kenyanthropus platyops (según Mauricio Antón). / Ilustración: Mauricio Antón
Más influyente todavía fue la propuesta de los paleontólogos Niles Eldredge y Stephen J. Gould de que buena parte de las secuencias evolutivas no se adecuaban al modelo simplista de cambio gradual, sino que las especies aparecían abruptamente durante momentos de cambio súbito y con posterioridad se mantenían estables hasta su extinción. En el campo de la evolución humana, el guante fue recogido por Niles Eldredge y Ian Tattersall, quienes en 1982 publicaron The myths of human evolution (“Los mitos de la evolución humana”), una obra que iba a tener una fuerte influencia en nuestra concepción de los patrones evolutivos en la evolución humana. A partir de ahí, el reconocimiento de que la evolución humana había sido un proceso mucho más variado y complejo se hizo notar en la interpretación de los antiguos fósiles y de otros nuevos que iban apareciendo. El panorama de la evolución humana cambió súbitamente y, bajo el nuevo paradigma, nuevas especies salieron a la palestra: Homo rudolfensis, Homo ergaster, Kenyanthropus platyops, Homo antecessor, Homo georgicus y otras más. Algunas especies anteriores saltaron literalmente por los aires, como el clásico Homo erectus, que dejó de ser un eslabón necesario en la evolución humana y quedó restringido al entorno asiático. Este paradigma mantuvo su vigencia durante los años ochenta y parte de los noventa del pasado siglo. A finales de esta década, sin embargo, se observan las primeras señales de desacuerdo por parte, por ejemplo, de Tim White, quien cuestiona la distinción entre Homo erectus y Homo ergaster. Pero son sobre todo los hallazgos en el yacimento georgiano de Dmanisi los que van a provocar de nuevo un cambio en el paradigma imperante.
El yacimiento de Dmanisi, datado por métodos radiométricos y de paleomagnetismo en 1,8 millones de años, ha proporcionado a lo largo de las últimas décadas el mejor registro fósil de los primeros homininos que salieron de África (Agustí y Lordkipanidze, 2005). Todo empezó en 1991, cuando el primer resto humano de Dmanisi, una mandíbula que conservaba toda su dentición, salió a la luz. Desde un principio esta mandíbula fue objeto de debate, ya que algunos autores cuestionaron su antigüedad en base al grado de reducción de los molares posteriores, y la situaron en el Pleistoceno medio y no en el Pleistoceno inferior. Hubo que esperar hasta 1999 para que el hallazgo de dos nuevos cráneos confirmase el carácter arcaico de los homininos de Dmanisi. El primero, D2280, solo conservaba la calota craneana, es decir, la parte más alta del cráneo que aloja el cerebro, lo que permitía inferir una capacidad craneana de 775 cm3. El segundo, D2282, muy deformado, conservaba además parte de la cara y su capacidad craneana pudo ser estimada en unos 650 cm3. En el año 2000 apareció una nueva mandíbula, esta vez de gran tamaño y con molares posteriores muy desarrollados, como correspondía a un representante arcaico del género Homo. Otros rasgos de la mandíbula evidenciaban un claro primitivismo, por lo que el paleontólogo georgiano Leo Gabunia y su equipo decidieron erigir una nueva especie, Homo georgicus. Esta mandíbula dio lugar a las primeras especulaciones relativas a la presencia de dos especies de homininos en Dmanisi.
Al año siguiente, en 2001, un nuevo cráneo acudió a la cita en Dmanisi. Esta vez, el cráneo D2700 incluía la mandíbula y se encontraba en un estado casi perfecto de conservación. El análisis detallado de este cráneo denotaba un hominino muy arcaico, con una capacidad craneana de 600 cm3. El perfil era cóncavo y, a diferencia del primer cráneo D2280, presentaba arcos superciliares poco desarrollados. La mandíbula era parecida a la primera mandíbula del año 1991 (Vekua et al., 2002). El cráneo D2700 demostró claramente que los homininos de Dmanisi no podían ser asignados a la especie africana Homo ergaster y que la primera salida de África fue protagonizada por poblaciones más próximas a Homo habilis que a esta última especie. Esta atribución fue confirmada en 2002, cuando salió a la luz un nuevo cráneo senil, un individuo desdentado (la mandíbula correspondiente apareció al año siguiente, en 2003) que, sin embargo, manifestaba los mismos rasgos arcaicos que el cráneo de 2001 (Lordkipanidze et al., 2005). La campaña del año 2005, finalmente, dio lugar al espectacular hallazgo del cráneo D4500, que encajaba con la gran mandíbula del año 2000. Este cráneo no defraudó las expectativas creadas por la mandíbula en su día. A pesar de que su capacidad craneana era la más baja de toda la población de Dmanisi, tan solo 546 cm3, mostraba una robustez que superaba el resto de anteriores hallazgos (Lordkipanidze et al., 2013).
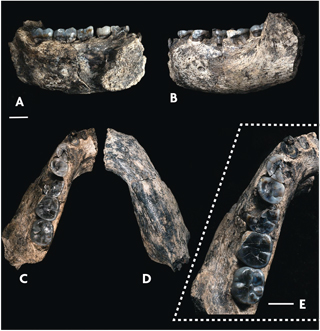
Vista del fósil de una mandíbula desde diferentes perspectivas. / Villmoare et al., 2015
El análisis del paleodeme completo de Dmanisi llevó al equipo paleoantropológico a la conclusión de que, pese a su variabilidad, el conjunto correspondía a una única población. El análisis geológico, paleomagnético y faunístico del yacimiento confirmaba este punto de vista: todos los restos de Dmanisi correspondían a un único episodio de sedimentación. Estos argumentos fueron contestados por aquellos que reconocían en el paleodeme de Dmanisi a más de una especie, quienes sin conocimiento de causa cuestionaron la estratigrafía del yacimiento. Para los autores del estudio del nuevo cráneo, sin embargo, no había duda: se trataba de una población arcaica de nuestro género que, a pesar de su limitada capacidad craneana, mostraba ya los caracteres derivados de Homo erectus.
La resistencia a aceptar que el conjunto de Dmanisi corresponde a una única población coetánea es comprensible, porque supone revisar buena parte de los criterios utilizados en África y otros enclaves para interpretar el registro fósil humano. La frontera entre Homo erectus y Homo ergaster, mantenida durante años, se diluye, lo mismo que la distinción entre Homo habilis y Homo rudolfensis. El caso es que, a diferencia de los múltiples hallazgos africanos, el paleodeme de Dmanisi corresponde a una única población que, con cinco cráneos y restos postcraneales de más de tres individuos, permite hacerse una idea de la variabilidad real de las primeras poblaciones del género Homo (Lordkipanidze et al., 2007).
La irrupción de la paleogenómica
Desde finales del siglo XX hasta la actualidad, nuestra interpretación del registro fósil se ha visto sacudida por la eclosión de un nuevo tipo de fósil, el ADN fósil, un tipo de registro inimaginable hace tan solo unas décadas. La secuenciación de ADN mitocondrial en organismos del Pleistoceno superior comenzó con éxito en la década de los ochenta del siglo XX y su aplicación a los neandertales, con cronologías que entraban dentro del rango de conservación de este material genético, estaba cantada. Curiosamente, entre los primeros restos neandertales en ser analizados, y que demostraron un aceptable nivel de conservación del ADN original, se encuentran los fósiles encontrados en el año 1857 en el valle de Neander y que sirvieron para bautizar al grupo. Con posterioridad, la muestra de neandertales en los que ha sido posible purificar auténtico ADN mitocondrial se ha ido ampliando, incluyendo los yacimientos de La Chapelle-aux-Saints en Francia, Engis en Bélgica, Mezmaiskaya en Rusia y Teshik Tash en Uzbekistán. Sin embargo, dos yacimientos –Vindija en Croacia y El Sidrón en España– destacan sobre el resto por el nivel de conservación de su ADN, por lo que han contribuido muy significativamente a la descodificación de esta molécula en los neandertales. En la actualidad, la recuperación de ADN mitocondrial ha podido extenderse hasta los ancestros de los neandertales, como es el caso del yacimiento de Sima de los Huesos en Atapuerca, cuyos restos humanos, atribuidos a Homo heidelbergensis, se remontan a cerca de 500.000 años.
Los primeros resultados sobre ADN mitocondrial en neandertales revelaron la existencia de secuencias muy diferentes a las del tipo humano actual (Krings et al., 1997). De un total de 379 pares de bases, se observaron 27 diferencias, que contrastan con las escasas 5 u 8 diferencias esperadas para una muestra al azar entre los humanos actuales. Ello significaba que la diferencia general entre las secuencias de los neandertales y los humanos modernos era tres veces superior a la media entre los humanos actuales, pero solo la mitad de la observada entre humanos y chimpancés. Si consideramos el tiempo de divergencia entre estas dos últimas especies, esto implicaba una notable antigüedad para la divergencia entre la línea que llevó al hombre anatómicamente moderno y la que desembocó en los neandertales: entre hace 550.000 y 690.000 años. El hecho de que las líneas que desembocaron en los dos grupos hubiesen evolucionado independientemente durante más de medio millón de años fue interpretado como una clara evidencia de que Homo neanderthalensis y Homo sapiens constituían dos especies diferentes.
Este resultado ponía punto final a un candente debate que se desarrolló a finales del siglo XX entre los partidarios del modelo multirregional, como Milford Wolpoff y Erik Trinkaus, y los partidarios del modelo «Arca de Noé», como Chris Stringer y Richard Klein, para explicar el origen del Homo sapiens. Según Wolpoff y Trinkaus, tras la primera salida de África, las diferentes poblaciones de Homo erectus habrían continuado manteniendo un elevado flujo genético, de manera que diferentes poblaciones regionales habrían desarrollado en paralelo las características de Homo sapiens. Frente a este modelo, la mayor parte de paleontólogos abogaban por un origen africano único de nuestra especie, previo a su dispersión por el resto de continentes. Pues bien, los resultados proporcionados por el análisis del mitogenoma neandertal confirmaban este último punto de vista al revelar que neandertales y humanos modernos constituían especies diferentes. El siguiente paso en la investigación fue el Proyecto Genoma Neandertal, abordado conjuntamente por parte de los equipos de Richard E. Green, de la Universidad de California, y Svante Pääbo, del Instituto Max Plank en Alemania. Esta vez ya no se trataba de reconstruir el ADN mitocondrial sino el genoma completo del neandertal, tal como se había hecho con el genoma humano anteriormente. Tras años de investigación, en los que de nuevo los yacimientos de Vindija y El Sidrón jugaron el papel más relevante, en 2010 se publicó un primer borrador que incluía aproximadamente un 60 % de la secuencia génica (Green et al., 2010) y llegaba a conclusiones que se confirmaron en 2013 con la publicación del genoma completo. Sorprendentemente, el análisis del ADN neandertal reveló la presencia de entre un 1 y un 4 % de secuencias neandertales en el genoma de los humanos modernos euroasiáticos, pero no en el genoma de las poblaciones africanas actuales. Ello quería decir que este limitado cruzamiento tuvo que haberse producido en los primeros momentos de la expansión de Homo sapiens fuera de África, muy probablemente cuando ambas poblaciones debieron encontrarse en el corredor de Palestina hace entre 40.000 y 60.000 años. Los multirregionalistas y los partidarios del cruce entre neandertales y humanos modernos acogieron este resultado con regocijo, ya que interpretaban que, de alguna manera, constituía un aval para sus ideas. En términos estrictos, este dato implicaba que, técnicamente, neandertales y sapiens no podrían considerarse especies diferentes.
Para la mayoría de paleoantropólogos, sin embargo, este resultado no modificaba significativamente la idea de que neandertales y humanos modernos representan en realidad especies o cuasi-especies diferentes, con historias evolutivas divergentes durante centenares de miles de años, sobre todo si se tiene en cuenta que, con posterioridad a su contacto en Palestina, ambas poblaciones volvieron a coexistir en Europa durante más de 10.000 años, sin que se volviesen a producir cruzamientos efectivos.
Pero el desvelamiento del genoma neandertal no fue el único resultado sorprendente aportado por la paleogenómica. El análisis realizado por el equipo de Pääbo de un pequeño fragmento de falange infantil procedente de niveles fechados entre 48.000 y 30.000 años de la cueva de Denísova, en los montes Altai en Siberia, reveló una secuencia de nucleótidos diferente no solo de la de los humanos actuales sino también de los neandertales (Krause et al., 2010). Los denisovanos, como así fueron llamados, habían habitado Asia central y habían llegado a cruzarse con los neandertales y con los humanos modernos. De hecho, entre las poblaciones modernas, el genoma denisovano mostraba las máximas concomitancias con los habitantes de Papúa Nueva Guinea. Sin embargo, no había rastro de genes denisovanos en poblaciones mucho más próximas, como los chinos.
Nos encontramos, por tanto, ante un hominino anterior a neandertales y humanos modernos, que llegó a convivir con ellos e incluso a cruzarse. Sin embargo, aparte de un par de molares de gran tamaño y la falange infantil, nada sabemos de este misterioso hominino. El estrecho parentesco con los neandertales apuntaría a poblaciones relictas de Homo heidelbergensis, pero podría tratarse de homininos todavía más arcaicos, tal vez poblaciones relictas de Homo erectus, que en un momento dado habrían llegado a cruzarse con humanos modernos en Australasia. En cualquier caso, la irrupción de los denisovanos ha acabado por romper los moldes en que se había basado la investigación paleoantropológica hasta la actualidad.
REFERENCIAS
Agustí, J., & Lordkipanidze, D. (2005). Los primeros pobladores de Europa. Barcelona: RBA.
Green, R. E., Krause, J., Briggs, A. W., Maricic, T., Stenzel, U., Kircher, M., ... Pääbo, S. (2010). A draft sequence of the Neandertal genome. Science, 328, 710–722. doi: 10.1126/science.-1188021
Krause, J., Fu, Q., Good, J. F., Viola, B., Shunkov, M. V., Derevianko, A. P., & Pääbo, S. (2010). The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature, 464, 894–897. doi: 10.1038/nature08976
Krings, M., Stone, A., Schmitz, R. W., Krainitzki, H., Stoneking, & Pääbo, S. (1997). Neandertal DNA sequences and the origin of modern humans. Cell, 90, 19–30. doi: 10.1016/S0092-8674(00)80310-4
Lordkipanidze, D., Jashashvili, T., Vekua, A., Ponce de León, M., Zollikofer, C., Rightmire, … Nioradze, M. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449, 305–310. doi: 10.1038/nature06134
Lordkipanidze, D., Ponce de León, M., Margvelashvili, A., Rak, Y., Rightmire, G. P., & Zollikofer, C. (2013). A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 342, 326–331. doi: 10.1126/science.1238484
Lordkipanidze, D., Vekua, A., Ferring, R., Rightmire, G. P., Agustí, J., Kiladze, G., … Zollikofer, C. (2005). The earliest toothless hominin skull. Nature, 434, 717–718. doi: 10.1038/434717b
Vekua, A., Lordkipanidze, D., Rightmire, G. P., Agustí, J., Ferring, R., Maisuradze, G., … Zollikofer, C. (2002). A new skull of early Homo from Dmanisi, Georgia. Science, 297, 85–89. doi: 10.1126/science.1072953
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, C. J., DiMaggio, E., Rowan, J., … Reed, K. E. 2015. Early Homo at 2.8 Ma from Ledi Geraru, Afar, Ethiopia. Science, 347, 1352–1355. doi: 10.1126/science.aaa1343




