

¿Me voy o me quedo?
Entendido el comportamiento de dispersión

Múltiples factores infl uyen en la decisión de dispersarse y en cuándo iniciar la dispersión, y su estudio proporciona evidencias de las causas y los determinantes de este proceso. Los animales abandonan el lugar de nacimiento para dar solución a tres problemas fundamentales: pérdida de calidad del hábitat, competencia por los recursos y riesgo de consanguinidad. La dispersión, sin embargo, no se lleva a cabo hasta que los individuos alcanzan una condición física conveniente, que les permita alcanzar con éxito un nuevo lugar de asentamiento. El proceso de dispersión tiene tres fases: salida del área natal, transición (desplazamiento) y asentamiento en el nuevo hábitat. Los individuos se enfrentan a diferentes decisiones en cada una de estas fases.
Palabras clave: calidad del hábitat, competencia, consanguinidad, dispersión, eficacia biológica, flujo génico.
«En prácticamente todas lass especies ocurre dispersión, y probablemente se trate de uno de los rasgos de historia de vida más importantes de los implicados en la evolución»
Uno de los conceptos más estudiados en ecología y biología evolutiva y, sin embargo, del que todavía queda mucho por comprender, es el de dispersión. La dispersión es el desplazamiento de individuos, o de estructuras especializadas para la dispersión (las diásporas, por por ejemplo, semillas), hasta un nuevo territorio donde tiene lugar al menos un episodio de reproducción. Al tratarse de desplazamiento, en los animales conviene evitar la posible confusión entre dispersión y migración. La migración se caracteriza por ser un desplazamiento periódico que implica a muchos individuos simultáneamente, y no siempre resulta en flujo génico (intercambio de variantes génicas) entre grupos o poblaciones. La dispersión, en cambio, implica a pocos individuos, y suele resultar en flujo génicos puesto que los individuos se incorporan a una nueva población en el seno de la cual se reproducen. Por ejemplo, las golondrinas se reproducen en toda la península Ibérica aprovechando granjas, establos y cortijos, donde conviven en grupos más o menos numerosos. A fi nales del verano realizan la migración hasta el África subsahariana, donde las condiciones para pasar el invierno son más favorables. Cuando a principios de la primavera regresan a sus zonas de reproducción, algunos individuos pueden decidir cambiar de emplazamiento y asentarse en otra granja o cortijo situado a unos metros o kilómetros de distancia de la anterior, donde podrán reproducirse con nuevas parejas potenciales. Podemos decir entonces que esos individuos han realizado migración estacional y luego, a su regreso para la reproducción, se han dispersado. La dispersión implica, por tanto, desplazamiento y flujo génico. Estas características hacen que se considere un rasgo con profundas implicaciones ecológicas y evolutivas para las especies. En prácticamente todas las especies se da la dispersión, y probablemente se trate de uno de los rasgos de historia de vida más importantes de los implicados en la evolución de las especies.
«Los individuos deben decidir en primer lugar si se dispersan o no, en segundo lugar cuándo iniciar la dispersión, y finalmente cuándo parar y establecerse»
Al abordar el estudio de la dispersión animal, las principales diferencias en el enfoque respecto al estudio de la dispersión en plantas tienen que ver con la capacidad de movimiento. En los animales son los propios individuos los que se desplazan, con esfuerzo locomotor, mientras que en las plantas el desplazamiento lo experimentan las diásporas, esto es, propágulos, frutos o semillas, y está mediado por factores ambientales (por ejemplo, viento o agua) o por animales. En los animales, además, puede darse dispersión antes de cada evento reproductor. Si bien la capacidad de movimiento constituye un rasgo fundamental que diferencia la dispersión en plantas y animales, resulta interesante comprobar que el patrón que muestran las distancias de dispersión en ambos grupos de organismos refleja una misma característica: el predominio de las distancias cortas respecto a grandes distancias (figura 1). De alguna manera, en ambos grupos los beneficios de la dispersión se alcanzan sin necesidad de grandes desplazamientos, que generalmente suponen un coste mayor.
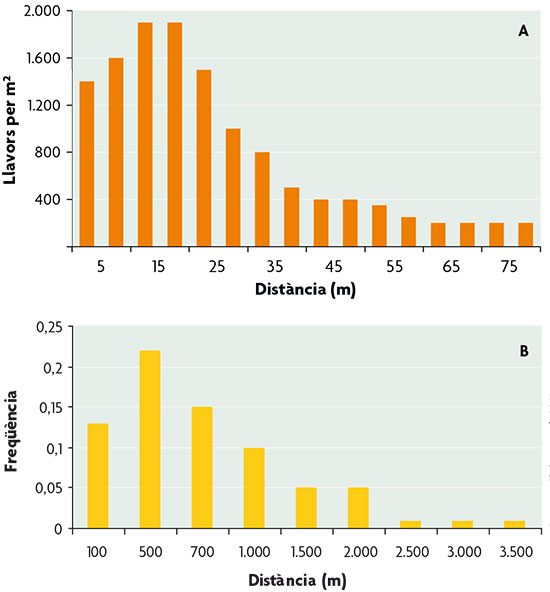
Figura 1. Distribución de las distancias de dispersión en una planta angiosperma (a: Alazán, Tachigalia versicolor) y en un ave paseriforme (b: Carbonero común, Parus major). / Basado en Clobert et al., 2001
Asociado a la capacidad de movimiento y al desplazamiento activo, otro factor clave en la dispersión animal es la capacidad de «decidir». Los individuos deben decidir en primer lugar si se dispersan o no, en segundo lugar cuándo iniciar la dispersión, y finalmente cuándo parar y establecerse. Estas mismas cuestiones existen en el caso de las plantas, pero se resuelven de la mano de los factores mediadores. Las consecuencias de la dispersión no son otras que las consecuencias del modo en que los organismos resuelven estas cuestiones, y los efectos sobre su eficacia biológica, es decir, sobre su supervivencia y reproducción, constituyen en última instancia las fuerzas evolutivas de la dispersión.
El proceso de dispersión tiene tres fases: salida del área natal, transición (desplazamiento) y asentamiento en el nuevo hábitat. Los animales se enfrentan a diferentes decisiones en cada una de estas fases. ¿Quiénes de los miembros de una población deciden dispersarse? ¿Por qué? ¿Cuándo iniciar la dispersión? ¿Cuándo parar? El estudio de la dispersión animal explora los factores que influyen en cada una de las decisiones. Una buena aproximación al resultado esperado en estas decisiones es la de considerar el balance de costes y beneficios de las decisiones posibles. ¿Dispersarse o quedarse? Múltiples factores influyen en la decisión de abandonar el área natal, y en algunos casos estos factores actúan de forma opuesta. Por ejemplo, los beneficios que proporcione alcanzar nuevos territorios actuarán a favor de la dispersión, mientras que el gasto energético o el riesgo de mortalidad durante el desplazamiento actuarán en contra.
«Frecuentemente los individuos que se dispersan no constituyen una muestra al azar, sino que exhiben una mayor eficiencia a la hora de desplazarse y colonizar nuevos territorios»
A través de los dilemas a los que se enfrentan los individuos, los biólogos nos asomamos a las grandes cuestiones que caracterizan la dispersión: las causas últimas (para qué sirve) y las causas próximas (qué factores la determinan). El estudio de la dispersión no resulta sencillo, porque requiere obtener medidas objetivas del desplazamiento y del éxito reproductor de los individuos dispersantes en los nuevos territorios. Gracias a los métodos demográficos y genéticos, el número de especies del que se tiene información va en aumento. El estudio científico de la dispersión en distintas especies animales proporciona evidencias de interesantes asociaciones entre variables relacionadas con la dispersión que difícilmente pueden considerarse meramente casuales, como veremos a continuación.
¿Quién se va? Características de los individuos que se dispersan
«Se considera que la dispersión puede cumplir tres funciones principales: mejorar la calidad de hábitat, evitar competencia intraespecífica y evitar consanguinidad»
Los individuos de una población difieren entre sí en sus características morfológicas, fisiológicas o comportamentales (es decir, tienen distintos fenotipos), mostrando por lo general una gran variabilidad intraespecífica. Entre ellos, frecuentemente los individuos que se dispersan no constituyen una muestra al azar, sino que exhiben una mayor eficiencia a la hora de desplazarse, colonizar nuevos territorios e integrarse en una nueva población. Es decir, se trata de fenotipos particulares que tienen mayor probabilidad de culminar con éxito la dispersión.
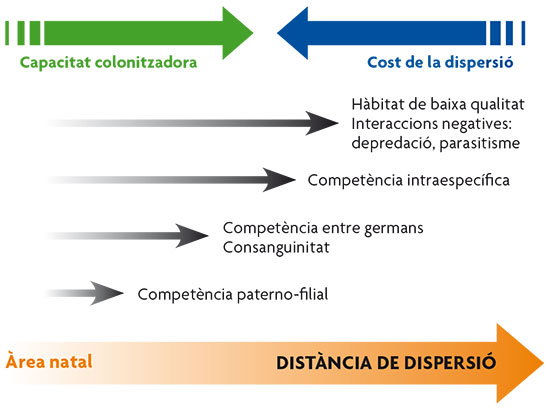
Figura 2. Esquema de las diferentes distancias de dispersión necesarias para escapar de las características negativas del área natal. / Basat en Clobert et al., 2001
Algunas especies proporcionan evidencias de diferencias morfológicas entre los individuos que se dispersan y los que no lo hacen. Por ejemplo, algunos insectos, como los pulgones (áfidos), presentan individuos alados y no alados, y la dispersión la realizan en una mayor proporción los individuos alados, que son los que pueden alcanzar mayores distancias. En otros casos los individuos que se dispersan difieren en rasgos comportamentales. Por ejemplo, en especies que viven en grupos sociales, la dispersión a un nuevo hábitat implica tener que integrarse en un grupo social nuevo, por lo que una predisposición a convivir con individuos que no pertenecen a su grupo propio puede ser beneficiosa. Es el caso de la rata-topo desnuda, un mamífero que vive en grupos sociales diferenciados. En ellos se ha comprobado que los individuos que se dispersan tienen un comportamiento diferente a los que no lo hacen: en experimentos donde se les ofrece aparearse con individuos de su propio grupo y con individuos de otro, tienen preferencia por los que no pertenecen a su grupo (O’Riain et al., 1996). Este comportamiento va a facilitar, sin duda, su integración en alguno de los grupos que encuentre en el nuevo hábitat.
¿Qué gana el que se va? Beneficios y costes de abandonar el territorio natal
Como se ha comentado anteriormente, la decisión de abandonar o no el lugar de nacimiento implica un balance de costes y beneficios. Los costes de la dispersión son numerosos, pues incluyen no solo el coste energético del desplazamiento y el riesgo de depredación sino el coste derivado del desconocimiento del nuevo hábitat, que implica, por ejemplo, buscar nuevas fuentes de recursos o nuevos grupos sociales para actividades de cooperación. También existe un coste por la pérdida de lo que conocemos como adaptación local, que hace referencia a aquellas sutiles adecuaciones a las condiciones ambientales concretas del hábitat de procedencia que habían llegado a desarrollarse y que se pierden en el nuevo hábitat. Es el caso de los ectoparásitos como las pulgas, que pueden llegar a desarrollar una alta adecuación a las condiciones concretas de temperatura que les ofrece el cuerpo del hospedador en que se han establecido (por ejemplo, un perro), de forma que la dispersión a un nuevo individuo hospedador puede acarrearles el coste de adaptarse a las nuevas condiciones locales. En general, los costes de la dispersión aumentan con la distancia recorrida, tal como refleja la figura 2.

Algunas especies proporcionan evidencias de diferencias morfológicas entre los individuos que se dispersan y los que no lo hacen. Por ejemplo, los pulgones presentan individuos alados y no alados, y la dispersión la realizan en una mayor proporción los individuos alados, que son los que pueden alcanzar mayores distancias. / David Martínez Torres
Por otra parte, se considera que la dispersión puede cumplir tres funciones principales (causas últimas): mejorar la calidad del hábitat, evitar competencia intraespecífica y evitar consanguinidad (Clobert et al., 2001). A continuación vamos a ver algunos ejemplos de evidencias a favor de cada uno de estos factores causales de la dispersión. No obstante, dependiendo de cuál de estas causas es la que ejerce una mayor presión, podemos esperar diferencias en el comportamiento de dispersión, por ejemplo, en las distancias recorridas (figura 2). Así, desplazarse una distancia corta, como puede ser moverse hasta el territorio adyacente, permite escapar de la competencia con los padres en animales territoriales. Distancias mayores serían necesarias para escapar de la competencia entre hermanos y del riesgo de consanguinidad, ya que ello implica, en muchos casos, cambiar de grupo social. Las distancias para colonizar territorios de mejor calidad se consideran de las más grandes que recorren los individuos.
En busca de El Dorado: cuando el objetivo es mejorar la calidad del hábitat
Existen interesantes evidencias que demuestran una relación entre dispersión y calidad del hábitat a partir de manipulaciones experimentales de algún componente relacionado con dicha calidad, como puede ser la presencia de especies con las que se producen interacciones negativas o la disponibilidad de recursos como alimento, refugio o lugares para la reproducción. Por ejemplo, un deterioro de la calidad del hábitat por el aumento de la presencia de depredadores en una población de pulgones mediante la introducción de uno de sus principales depredadores, las mariquitas, provoca el nacimiento de una mayor proporción de individuos alados en la siguiente generación (Weisser et al., 1999). A la inversa, la eliminación de parásitos en nidos de aves, una clara mejoría de ese hábitat reproductor, retrasa el inicio de la dispersión de los polluelos.
En cuanto a los recursos, una mayor disponibilidad de alimento se ha visto que puede determinar la disminución de la dispersión. Así ocurre con las gaviotas, que ante el incremento de la presencia de vertederos en las áreas costeras actualmente han relajado sus distancias de dispersión. También la existencia de los descartes de pescado practicados por los barcos de pesca ha hecho que aves marinas como las pardelas hayan aprendido la existencia de esa fuente de recurso diario, lo que ha reducido la dispersión hacia zonas donde no hay actividad pesquera. La disponibilidad de refugios se ha visto que también constituye un factor determinante de la ocupación de un territorio, por ejemplo para los reptiles. Así, la creación de refugios artificiales mediante la colocación de troncos puede incrementar notablemente la llegada de reptiles a un área. Lo mismo ocurre con la disponibilidad de lugares para la reproducción, que puede facilitar la dispersión de algunas aves hacia los lugares donde se ha incrementado experimentalmente la disponibilidad de estos sitios mediante la colocación de cajas-nido.

En los animales conviene evitar la confusión entre dispersión y migración. Las golondrinas se reproducen en toda la península Ibérica aprovechando granjas, establos y cortijos, donde conviven en grupos más o menos numerosos. A finales del verano realizan la migración hasta África. Cuando a principios de la primavera regresan a sus zonas de reproducción, algunos individuos se dispersan: pueden decidir cambiar de emplazamiento y asentarse en otra granja o cortijo, situada a unos metros o kilómetros de distancia de la anterior. / Dori
Cuando la familia es el problema
Algunos estudios experimentales proporcionan evidencias de la influencia de la competencia entre individuos de la misma especie en la dispersión. Por ejemplo, un aumento experimental en la densidad de individuos aumenta la dispersión en numerosas especies de insectos, mientras que un aumento experimental en la cantidad de alimento disminuye la dispersión.
Numerosas evidencias proceden del particular tipo de competencia intraespecífica entre individuos emparentados, esto es, entre hermanos, y entre padres e hijos. Por ejemplo, para evitar la competencia entre hermanos, algunos padres fuerzan la marcha de una fracción de su descendencia, incluso aunque la dispersión conlleve la mortalidad de algunos individuos. Esto se ha observado en algunas aves, como los arrendajos, donde el beneficio en eficacia biológica suele ser mayor para los padres (que aseguran la supervivencia de la fracción que se queda) que para el individuo que se dispersa. Otras evidencias de que la dispersión puede responder a la competencia entre hermanos se produce en especies con interacciones entre hermanos dominantes y subordinados, donde el resultado es la marcha del subordinado. Una evidencia indirecta de la importancia de esta competencia procede de algunas especies de mamíferos, como el zorro, donde a menores tamaños de camadas se observa una menor tendencia a la dispersión.
La competencia entre padres e hijos suele darse en especies que dependen fuertemente de un territorio para desarrollar su actividad. En estas especies, la adquisición del territorio por parte de los juveniles puede empezar pronto, entrando así en conflicto, aparte de con los hermanos, con los padres. En algunos casos, como ocurre con la ardilla de tierra, los padres fuerzan la dispersión solo de las crías adelantadas (ya que tienen mayor probabilidad de adquirir un territorio que las retrasadas); en otros casos, como en el gorrión cantor, son las crías las que expulsan agresivamente a los padres del territorio; en un tercer grupo de casos, como el que se da en la ardilla roja, son los padres los que abandonan el territorio antes de entrar en competencia por él con las crías. Una evidencia indirecta de que la competencia entre individuos emparentados favorece la dispersión es que la existencia de cooperación entre ellos hace que esta disminuya, como ocurre en los leones y en algunas especies de primates.

En el caso de la rata-topo desnuda, los individuos que se dispersan tienen un comportamiento diferente a los que no lo hacen: muestran una preferencia por los que no pertenecen a su grupo. Este comportamiento va a facilitarles la integración en alguno de los grupos que encuentren en el nuevo hábitat. / Kevin McGee
Cuando el incesto es el peligro: evitar los riesgos de la consanguinidad
En numerosas especies la dispersión está sesgada a uno de los dos sexos, lo que parece una forma perfecta de evitar la consanguinidad. Curiosamente, en unas especies son los machos los más propensos a dispersarse y en otras son las hembras. Las evidencias llevan a explicar cada caso en función de los costes y beneficios de la dispersión para cada sexo, de forma que la dispersión de uno de los dos sexos disminuye la probabilidad de aparearse con parientes al tiempo que proporciona beneficios específicos al sexo que se va. Estos beneficios se ven influidos por el sistema de apareamiento y por la estructura social que caracteriza a la especie. Respecto al sistema de apareamiento, cuando se trata de monogamia (machos que se aparean con una sola hembra) es más frecuente la dispersión de las hembras, mientras que en poliginia (machos que se aparean con varias hembras) es más frecuente la dispersión de los machos. Respecto a la estructura social, cuando es la defensa de recursos la que determina la estructura social, aumenta la dispersión de hembras, mientras que cuando es la defensa de las hembras por parte de los machos lo que constituye la base de la estructura social, aumenta la dispersión de los machos.
«Para evitar la competencia entre hermanos, algunos padres fuerzan la marcha de una fracción de su descendencia, incluso aunque la dispersión conlleve la mortalidad»
Las aves y los mamíferos constituyen los grupos que aportan más evidencias de la importancia de estos factores. En las aves, los machos compiten por el territorio, por lo que se benefician más que las hembras de la familiaridad con este. Por ello, los machos se dispersan menos que las hembras. Estas, en cambio, eligen al macho atendiendo al territorio que posee (o al nido que construye), y la dispersión les permite elegir entre un mayor número de territorios o nidos ofrecidos por los machos. Por ello, las hembras se dispersan más que los machos. Por el contrario, en los mamíferos, son las hembras las que compiten por el territorio y se benefician más que los machos de la familiaridad con este. Ello hace que se dispersen menos que los machos (Cockburn et al., 1985). Estos, en cambio, eligen el territorio por la presencia en él de hembras, y se benefician de la dispersión al poder acceder a un mayor número de territorios con hembras. Por ello, los machos se dispersan más que las hembras.

En juveniles de lagartijas, los niveles de corticosterona –una hormona relacionada con la actividad locomotora y la respuesta ante el estrés– están relacionados con su propensión a la dispersión. / Jacinta Lluch Valero
Dispersarse sí, pero ¿cuándo iniciar la dispersión?
La dispersión está determinada por tres grupos de factores (causas próximas): factores ambientales, factores internos de los individuos que se desplazan y factores genéticos.
Las condiciones ambientales constituyen una de las causas de la dispersión. No obstante, los animales no siempre responden directamente a las condiciones del ambiente, sino que en algunos casos la respuesta es indirecta, mediada por un efecto previo en la condición física de los individuos. En general, cambios rápidos e impredecibles en las condiciones ambientales, como por ejemplo invasiones de enemigos naturales, poca disponibilidad de parejas o episodios climáticos extremos, llevan a una respuesta directa de dispersión. Sin embargo, cambios graduales y predecibles en las condiciones ambientales, como por ejemplo estacionalidad o disponibilidad de recursos, resultan en una respuesta indirecta de dispersión.
Dos tipos de factores internos determinan el inicio de la dispersión: la condición física de los individuos y los niveles de determinadas hormonas en la sangre. Tener una condición física robusta parece ser un requisito previo al inicio de la dispersión. Las evidencias indican que algunos individuos retrasan la partida del área natal hasta que las reservas energéticas almacenadas son las adecuadas para resistir la dispersión. Asimismo, tener niveles altos de las hormonas relacionadas con la capacidad de mostrar agresividad, como es el caso de la testosterona, y con la actividad locomotora y la respuesta frente a situaciones de estrés, como es el caso de la corticosterona, parece también un requisito para iniciar la dispersión. Existen evidencias interesantes en mamíferos, aves y reptiles sobre el papel de estas hormonas en la dispersión (Belliure y Clobert, 2004). Por ejemplo, embriones de mamíferos como los ratones, expuestos a testosterona durante la gestación, tienen una mayor predisposición a la dispersión. En las aves, el nivel de testosterona presente en los huevos influye en la capacidad de dispersión del polluelo al nacer, que es mayor en los de mayor nivel de testosterona. En juveniles de lagartija, los niveles de corticosterona están relacionados con la capacidad locomotora y la propensión a la dispersión.

Una mayor disponibilidad de alimento puede determinar la disminución de la dispersión. Así ocurre con las gaviotas que ante el incremento de la presencia de vertederos en las áreas costeras actualmente han relajado sus distancias de dispersión. / Grand Parc de Burdeos
Por otra parte, preguntarse por la base genética de la dispersión es cuestionarse si existe variación genética en los caracteres relacionados con la dispersión y si los caracteres implicados en la dispersión se expresan siempre juntos, es decir, si están correlacionados genéticamente. Las evidencias indican la existencia, al menos en algunas especies animales, de ambas componentes de la determinación genética. Por ejemplo, en el caso de los pulgones existe variación genética entre los individuos alados y no alados. Y en algunas especies de insectos ortópteros como los grillos, existe evidencia de correlación genética entre tres caracteres relacionados con la dispersión, como son la propensión al vuelo, la forma del ala y la fisiología muscular para generar energía, tres rasgos implicados en la dispersión de los grillos que se expresan siempre de forma conjunta.
En definitiva, y como individuos pertenecientes a una especie animal que también proporciona muchas evidencias de lo que acabamos de exponer, abandonar o no el lugar de nacimiento constituye una de las preguntas clave que afrontan, o afrontamos, la mayoría de organismos. Ambas opciones tienen sentido en el contexto de diferentes escenarios ambientales, y en la adecuación de las respuestas basa su éxito la vida. Los factores que influyen en la dispersión se convierten en potentes fuerzas evolutivas. El estudio de la dispersión animal, sin duda, continuará proporcionando asombrosas evidencias del poder de dichas fuerzas evolutivas para moldear un rasgo tan complejo y con tan profundas consecuencias para las especies como el que constituye el comportamiento de dispersión.
Belliure, J. i J. Clobert, 2004. «Behavioral Sensitivity to Corticosterone in Juveniles of The Wall Lizard». Physiology and Behavior, 81: 121-127. DOI: <10.1016/j.physbeh.2004.01.008>.
Clobert, J.; Danchin, E.; Dhondt, A. A. i J. D. Nichols (eds.), 2001. Dispersal. Oxford University press. Oxford.
Cockburn, A.; Scott, M. P. i D. J. Scotts, 1985. «Inbreeding Avoidance and Male-Biased Natal Dispersal in Antechinus spp. (Marsupialia: Dasyuridae)». Animal Behaviour, 33(3): 908-915. DOI: <10.1016/S0003-3472(85)80025-7>.
O’Riain, M. J.; Jarvis, J. U. M. i C. G. Faulkes, 1996. «A Dispersive Morph in The Naked Mole-rat». Nature, 380: 619-621. DOI: <10.1038/380619a0>.
Weisser, W. W; Braendle, C. i N. Minoretti, 1999. «Predator-induced Morphological Shift in The Pea Aphid». Proceedings of the Royal Society London B, 266: 1175-1181. DOI: <10.1098/rspb.1999.0760>.




