

Reflexiones sobre el origen de la vida
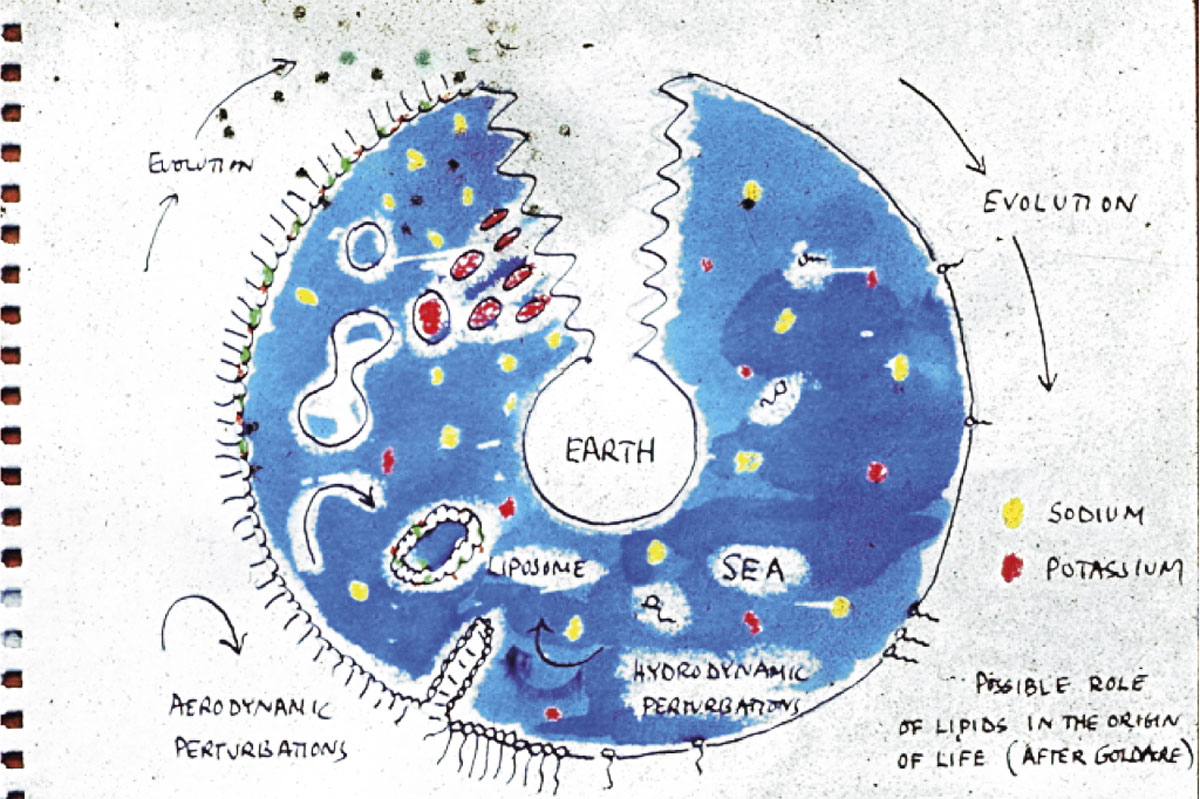
En este artículo argumentamos que el problema del origen de la vida no se puede explicar apelando exclusivamente a los mecanismos de la evolución darwiniana, como un gran número de expertos tiende a asumir, sino que requiere un profundo cambio de perspectiva. En esa línea, destacamos que la selección natural, para operar como motor de diversificación (e, indirectamente, de potencial incremento de la complejidad), implica un conjunto de condiciones que, de hecho, la hagan posible: más en concreto, organizaciones químicas automantenidas y auto(re)productivas con un espacio fenotípico (es decir, un abanico de funciones) lo suficientemente amplio. Así pues, como alternativa a los planteamientos mayoritarios en el campo del origen de vida, basados en poblaciones moleculares (habitualmente de ARN) sujetas a evolución darwiniana, proponemos una extensión del paradigma «autoorganizativo» hacia uno «auto(re)productivo», que recoja adecuadamente la especificidad del fenómeno biológico (en particular, su dimensión celular y metabólica), y que tendría relevancia antes, durante y después de que la selección natural comenzara a operar.
Palabras clave: autoorganización, protocélulas, metabolismo, diversidad fenotípica/funcional, evolución abierta.
Muchos científicos y filósofos comparten actualmente la idea de que el origen de los seres vivos a partir de la materia inerte, aún siendo una problemática complicada y todavía sin terminar de cerrar, se encuentra, digamos, «en vías de solución». Obviamente queda un gran número de «detalles» por resolver, que sin duda pueden ser difíciles cuestiones para los especialistas del campo; pero, desde una perspectiva global, se asume que podemos llegar a entender esta transición fundamental, gracias a los resultados de los experimentos de Miller y sus diversas secuelas (es decir, al campo de la «química prebiótica» que ha logrado la síntesis de diversos compuestos relevantes para la vida) y a la teoría de la evolución, aplicada a las primeras moléculas replicativas (nucleótidos de ARN o análogos). Sin embargo, algunas voces críticas han puesto en cuestión esta optimista visión. Por ejemplo, en su reciente libro Mind and Cosmos. Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False, el filósofo Thomas Nagel (2012) sostiene que las ciencias biológicas contemporáneas son incapaces de explicar el origen de la vida, o el origen de la mente. Más concretamente, Nagel afirma que ninguna teoría científica puede proporcionar una explicación satisfactoria –lo que para él significa una explicación reduccionista a las ciencias físicas– de cómo podrían haber aparecido fenómenos tan complejos a partir de la evolución del mundo físico-químico. Nagel en su libro tiende a identificar el sentido explicativo del término reduccionismo como equivalente a materialismo reduccionista, porque no considera que las teorías «emergentistas» ofrezcan una explicación alternativa de este tipo de sistemas de alta complejidad o de sus respectivas propiedades, biológicas y cognitivas. Como él mismo dice: «Que tales elementos puramente físicos, cuando son combinados de cierta forma, deban producir necesariamente un estado del todo que no se constituye a partir de las propiedades y las relaciones atribuibles a sus partes físicas, sigue sonándome a magia».
«Una consideración detenida sobre las diferencias entre la materia viva y la no viva debe llevarnos a reflexionar más sobre las dificultades que encierra el problema del origen de la vida»
Y es que, efectivamente, una consideración detenida sobre las diferencias entre la materia viva y la no viva debe llevarnos a reflexionar más sobre las dificultades que encierra el problema del origen de la vida. Explicar la transición entre el mundo físico-químico y el biológico es un reto de enormes dimensiones, principalmente por el profundo contraste existente entre ambos dominios hoy en día. Cualquier ser vivo conocido, incluso unicelular, es órdenes de magnitud más complejo (en composición, diversidad de interacciones, comportamiento dinámico) que los distintos sistemas moleculares de los cuales se ha postulado que pudiera provenir. Cuando analizamos los componentes, las transformaciones químicas, los mecanismos de control que subyacen a las propiedades de las células más sencillas sobre nuestro planeta (procariontes con genomas muy reducidos), resulta imposible entender, de una sola vez, todo lo que está ocurriendo en su interior y en su relación con el entorno. Innumerables reacciones de síntesis (bio)química, junto a procesos de transporte, transducción, reconocimiento, señalización… ocurren cada segundo en la forma necesaria para que el organismo se mantenga, se adapte y, potencialmente, se reproduzca. Esto es posible gracias a una batería de mecanismos moleculares que los propios seres vivos han generado (en particular, mecanismos metabólicos, de síntesis autónoma y control regulativo [Ruiz-Mirazo y Moreno, 2012]), que los hacen especialmente robustos, pese a ser sistemas abiertos y alejados del equilibrio termodinámico. Ningún otro tipo de sistema material conocido, natural o artificial, exhibe, ni de lejos, niveles comparables de complejidad, tanto en un sentido estructural como organizativo o funcional.
El contraste entre ambos mundos resulta muy evidente al examinar cada ser vivo individualmente, pero también cuando consideramos el fenómeno biológico desde una perspectiva temporal más amplia. A escalas de tiempo mucho mayores que las fisiológicas, la vida se nos manifiesta como evolución, es decir, como un encadenamiento histórico entre organizaciones o entidades efímeras, que sin embargo trasmiten a sus sucesoras sus rasgos específicos. Es verdad que, si ampliamos el marco lo suficiente, todo (incluso un átomo, una ley física) puede entenderse como fruto de un proceso de transformación, de evolución temporal. Sin embargo, una vez que las piezas y las reglas de interacción básicas quedaron establecidas, la materia ha implementado modos de evolución a largo plazo sorprendentemente diferentes. Por un lado, la materia inanimada conocida se ha regido de manera estricta por las leyes fundamentales que caracterizan nuestro universo físico en expansión, generando estrellas, planetas, discos de asteroides, polvo interestelar, galaxias, y agujeros negros. Por otro, la materia que ha integrado (aunque sea sólo local y temporalmente) los sistemas biológicos ha contribuido al desarrollo de mecanismos de control sobre su variación mucho más sofisticados e indirectos, pese a ser plenamente congruentes con las leyes fisicoquímicas.
Estos mecanismos moleculares de control de la variación, partiendo curiosamente de condiciones de estabilidad precaria (estados estacionarios alejados del equilibrio termodinámico), se fueron fijando de un modo cada vez más consistente en diversas poblaciones de protoorganismos, a los cuales confirieron mayor robustez y adaptabilidad metabólica. Así, consiguieron transformarse, a lo largo del proceso de origen de vida, en módulos cada vez más complejos y, al mismo tiempo, de mayor perdurabilidad, órdenes de magnitud mayor que el tiempo de vida medio de esos protoorganismos de los que, de hecho, provenían. Al cabo, ciertos polímeros proclives a copia y conservación de su secuencia pasaron a convertirse en «registros moleculares» (Pattee, 1969) parciales, pero muy fiables, de la complejidad estructural y organizativa de las células en las que se fueron alojando y dentro de las cuales han ido tomando cuerpo y sentido desde entonces hasta la fecha. Fue precisamente la emergencia y funcionalidad de estos «componentes registro», con capacidad de replicación y transferencia fiable a otros sistemas celulares similares, lo que permitió que la vida lograra establecerse como un fenómeno sostenible a largo plazo sobre la superficie de la Tierra, gracias a una forma de evolución de la materia que había sido inaccesible hasta ese momento: la evolución darwiniana (Lewontin, 1970) o evolución abierta (Ruiz-Mirazo y Moreno, 2012). Por asombroso que pueda parecer, actualmente contamos con evidencia muy sólida de que este modo de cambiar y reinventarse que caracteriza a lo vivo, basado en poblaciones de metabolismos genéticamente instruidos, lleva en marcha, al menos sobre nuestro planeta, varios miles de millones de años.
La selección natural no explica el origen
La importancia del proceso evolutivo biológico y su formidable profundidad temporal, como nos mostró Darwin –y como toda la biología posterior a él ha apuntalado– ha llevado a otorgar a estos mecanismos de control de la variedad molecular (los mecanismos genéticos, hereditarios) un papel fundamental en el despliegue del dominio de lo vivo sobre la Tierra. Esto, junto a la falta de una teoría adecuada sobre la organización biológica (sobre la que volveremos enseguida), ha generado un importante desequilibrio que queda plasmado en concepciones probablemente erróneas –o cuando menos fuertemente descompensadas– sobre el problema del origen de vida. Nos referimos, en particular, a la asunción de que la teoría de la evolución (y, más específicamente, el mecanismo de «selección natural» aplicado a poblaciones de moléculas autorreplicantes, como ARN) sea la clave para resolver la transición entre el dominio de lo inerte y el de lo vivo. Esta asunción, ampliamente aceptada en círculos científicos desde hace varias décadas (Eigen y Schuster, 1979; Orgel, 1992; Szostak, 2012), es paradójicamente contraria a la intuición del propio Darwin, quien, a pesar de tener una visión del origen muy avanzada (similar en muchos aspectos a la que se fraguó un siglo más tarde [Peretó, Bada y Lazcano, 2009]), consideraba a la ciencia de su tiempo, incluida la teoría sobre la evolución que él mismo estaba impulsando en esos momentos, limitada para poder encarar dicha problemática.
«Ha sido una interpretación simplista del legado de Darwin lo que ha llevado a considerar la selección natural como la principal –para algunos incluso la única– fuerza o guía en el proceso de emergencia de vida»
En realidad ha sido una interpretación simplista del legado de Darwin lo que ha llevado a considerar la selección natural como la principal –para algunos incluso la única– «fuerza» o «guía» en el proceso de emergencia de vida. No obstante, como argumentamos en más detalle en Moreno y Ruiz-Mirazo (2009), apoyándonos en autores como Lewontin (1970) o Wicken (1987), entre otros, el establecimiento de un modo de evolución darwiniano, que realmente opere por selección natural, requiere como condición de partida un espacio fenotípico lo suficientemente amplio, es decir, un potencial de variación y diversificación funcional que resulta difícilmente alcanzable para formas de organización material por debajo del umbral de complejidad asimilable al de protocélulas con metabolismo propio. En otras palabras, el mecanismo de evolución por selección natural no sirve para explicar el origen de la vida porque, en el fondo, la selección natural implica ya la existencia de organismos, o «protoorganismos» con un fenotipo (o un conjunto de funciones atribuibles a sus componentes) lo suficientemente rico. Entonces, ¿dónde buscar principios o mecanismos que expliquen cómo, en las condiciones de la Tierra primitiva, haya podido generarse un proceso sostenido de cambios que llevara a la aparición de la vida? ¿Hay alguna ley o principio en el mundo físico que permite concebir la aparición de la vida como una necesidad o debemos resignarnos a pensar, como nos decía Monod (1970), que el origen de la vida es tan improbable, tan fruto del azar, que se trata prácticamente de un milagro? ¿Cómo puede la materia originar algo que parece ser tan profundamente diferente de sus propiedades iniciales?
¿En qué consiste el cambio de perspectiva que proponemos?
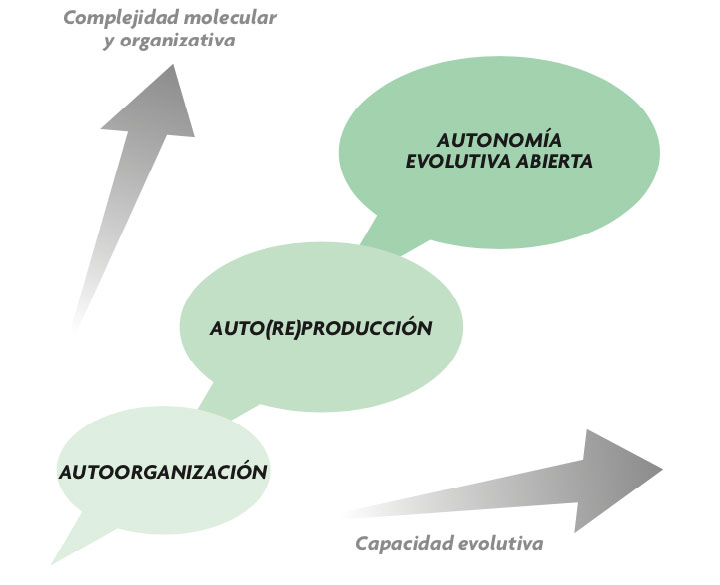
Una premisa de la que podemos partir, con garantías de consenso pleno en la comunidad científica, es que la naturaleza no da saltos en el vacío y, por tanto, se debe plantear un continuo entre la materia no viviente y la vida. Es decir, debe haber algún tipo de explicación progresiva, en etapas de creciente complejidad (véase figura 1), de la transición hacia la vida, aunque en el límite nos podamos ver abocados a una narrativa histórica, en gran medida contingente, del proceso. Como Fry (2000) ha señalado, el problema fundamental del origen de la vida radica en la tensión entre el principio de continuidad y la dificultad de explicar las diferencias obvias entre la no-vida y la materia viva. Si el origen de la vida es una cuestión científica legítima (y creemos que lo es), uno debe buscar una teoría que sirva de puente entre la física y la biología, y demostrar experimentalmente que ese puente (al menos una parte importante del mismo) se puede reconstruir de novo. A fin de cuentas, los seres vivos están hechos de los mismos elementos materiales que los sistemas no vivos, así que no parece descabellado tratar de esclarecer el vínculo que debió establecerse entre unos y otros.
«La naturaleza no da saltos en el vacío y, por tanto, se debe plantear un continuo entre la materia no viviente y la vida»
Desde la perspectiva de las ciencias físicas, explicar el origen de la vida es una tarea muy difícil porque cuanto más complejo es un sistema, menos probable resulta su aparición y persistencia. En primera aproximación, parece más o menos fácil de entender cómo agregados materiales simples pueden generar espontáneamente estructuras compuestas relativamente estables (moléculas complejas, macromoléculas o agregados supramoleculares) en virtud de los diferentes tipos y grados de fuerza que se dan en la naturaleza (Simon, 1962): efectivamente, como resultado de estas interacciones físicas de distinta magnitud, es previsible la formación de estructuras más complejas (con propiedades que en muchos casos se podrían considerar «emergentes», no presentes en sus partes separadas –como por ejemplo, la superconductividad, la afinidad química, o la topología cerrada de una vesícula lipídica). Pero a medida que la complejidad de las estructuras crece su mantenimiento se convierte en un problema: el ruido térmico aumenta la fragilidad y, por otra parte, la coincidencia o la coordinación de muchos procesos altamente específicos se convierte en algo cada vez más improbable. Además, la complejidad de la vida no sólo reside en la estructura de sus componentes, sino en una intrincada red de interacciones dinámicas que se establecen en condiciones alejadas del equilibrio termodinámico. Dicho de otra manera, estamos obligados a encarar el problema en términos organizativos. Los sistemas biológicos, de hecho, hacen uso de las propiedades autoorganizativas de la materia en muy diversas formas (Karsenti, 2008), unas más alejadas del equilibrio que otras. Y gracias a los avances que se dieron durante la segunda mitad del siglo xx en química supramolecular y en termodinámica de procesos irreversibles, contamos con un marco adecuado para entender la formación de organizaciones dinámicas relativamente complejas, que podrían llegar a combinar distintos agregados supramoleculares y patrones de orden de tipo «estructura disipativa».
Sin embargo, la vida va más allá: no sólo organiza componentes materiales preexistentes, sino que genera y regenera continuamente la mayoría de ellos y, al tiempo, pasa a controlar buena parte de las condiciones en las que dicha organización dinámica tiene lugar. Es decir, todo sistema biológico construye una organización dinámica a través de procesos de síntesis y transformación de componentes, incluyendo aquellos que procuran sus propias condiciones de viabilidad y mantenimiento en el tiempo. Las implicaciones de este paso desde la autoorganización a la autoproducción son múltiples, pero se pueden resumir en dos (Ruiz-Mirazo y Moreno, 2012): (i) un tipo de cohesión interna y robustez dinámica mucho mayor, basada en la «integración funcional» de los componentes del sistema; y (ii) un incipiente control sobre las condiciones de contorno bajo las cuales dicho sistema opera, es decir, la aparición de la «agencialidad». Todo esto está relacionado, en suma, con la emergencia de los sistemas metabólicos: una problemática que pone en evidencia que si aspiramos a elaborar una teoría general, sólida y congruente, sobre la organización biológica, ésta debe tomar en consideración lo aportado por las ciencias de la complejidad y el paradigma de la autoorganización, pero, sin duda, sobrepasarlo, acercándose de un modo más específico a la fenomenología de lo vivo (Keller, 2007).
«Desde la perspectiva de las ciencias físicas, explicar el origen de la vida es una tarea muy difícil porque cuanto más complejo es un sistema, menos probable resulta su aparición y persistencia»
Desde esta perspectiva habría que centrarse, entonces, en investigar distintas combinaciones de componentes moleculares y procesos de interacción y transformación, hasta dar con algunas que (siendo prebióticamente plausibles) consiguieran una mayor cohesión, un mantenimiento dinámico más estable, como condición de posibilidad para ulteriores transiciones. Consideramos, de hecho, muy razonable plantear que en ciertos entornos de la superficie terrestre hace unos 3.500 millones de años debieron formarse, impulsados por fuentes de energía externa (como el sol o la energía geotérmica), múltiples conjuntos de reacciones químicas alejadas del equilibrio en las que se producían gran diversidad de compuestos, más o menos estables (figura 2). En este contexto, también es muy razonable asumir que parte de estos compuestos o agregados de compuestos, si se acumulan lo suficiente (sobre una superficie absorbente o en un microcompartimento), tendrán un efecto sobre compuestos y procesos adyacentes, de tal manera que potencialmente contribuyan al mantenimiento de todo el conjunto de reacciones (del mismo modo que, en un ciclo autocatalítico sencillo, cada componente del ciclo contribuye a la síntesis del resto). La idea es que esas formas de cohesión y automantenimiento robusto se lograrían cuando las influencias entre los distintos componentes en interacción tuvieran un efecto de refuerzo mutuo, evitando tendencias naturales como la dispersión espacial o el decaimiento de estructuras materiales de no equilibrio. No resulta sencillo determinar el conjunto mínimo de componentes o procesos necesario para obtener una integración funcional autónoma, y por ello es una cuestión que se debe investigar empíricamente. De todas formas, la presencia de mecanismos de control cinético (catalizadores) y espacial (compartimentos), así como su adecuada coordinación, parece un requisito indispensable.
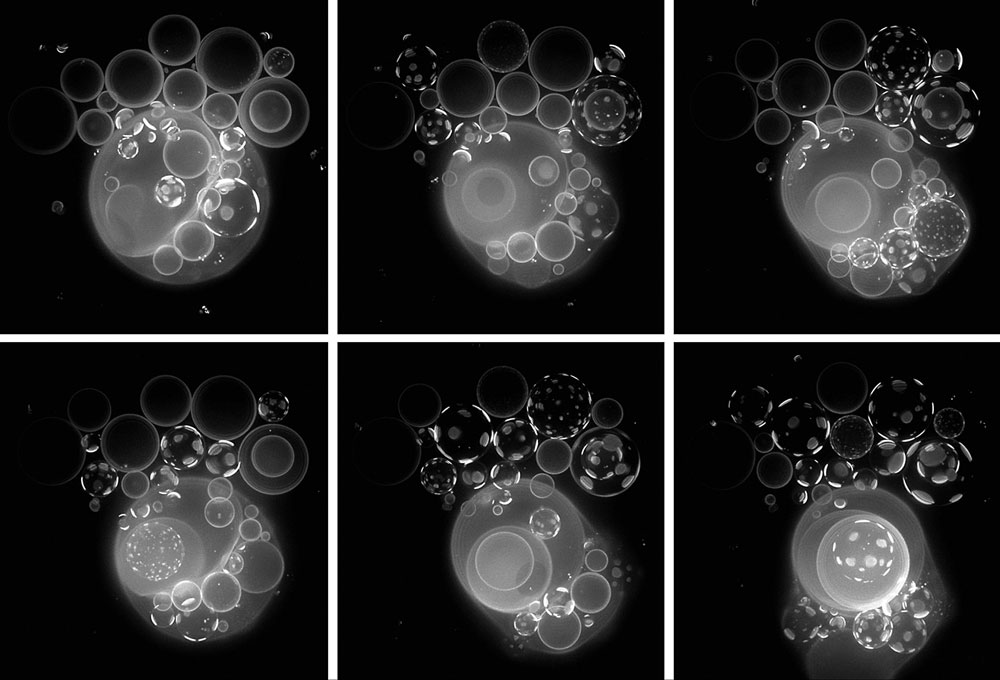
En cualquier caso, lo interesante es que estos sistemas no sólo serían viables en virtud de unas determinadas condiciones externas (las cuales siempre podrían hacerlos desaparecer, si fueran lo suficientemente adversas). Ellos mismos crearían y mantendrían gran parte de las condiciones que de hecho posibilitan su constitución y su progresión dinámica, ya que es la presencia de los diversos componentes y su implicación en el conjunto de procesos de interacción y transformación lo que hace posible la realización del resto. Estamos ahora en un escenario donde opera un régimen causal radicalmente nuevo: grupos de moléculas generan un conjunto de patrones y estructuras materiales (otras moléculas o agregados de moléculas, como cadenas peptídicas con capacidad catalítica o compartimentos lipídicos de permeabilidad selectiva –véase figura 3) que constriñen los procesos y transformaciones subyacentes de modo que regeneran recursivamente dichas estructuras y, a partir de ahí, el conjunto de las interacciones que mantienen dinámicamente todo el sistema. Esta es una idea que otros autores (Kauffman y Clayton, 2006) han plasmado en términos de «ciclos de constricción-trabajo», sugiriendo que las constricciones aparecen y se propagan siempre que alguna configuración material en una parte del universo establece bucles no lineales y recursivos de este tipo, y que es un fenómeno clave para entender el origen de la vida. Se trata, pues, de armar un entramado causal que, lejos de restringir o acotar el espacio de posibles estados dinámicos del sistema, permita la existencia de nuevos estados estacionarios estables, reflejando la mayor cohesión lograda entre sus componentes.
Implicaciones del enfoque alternativo
Llegado este punto, estamos ya en condiciones de responder al desafío planteado por Nagel. La materia ha podido transitar del universo fisicoquímico al biológico al encontrar, como resultado quizá también de eventos contingentes, formas estables de organización dinámica que surgen de su capacidad de «autoconstricción». Estas organizaciones, basadas en un conjunto de bucles de interacción recursiva, no lineal, no son exclusivas de la biología: en el mundo de los fenómenos físicos han aparecido y aparecen espontáneamente, como vemos en el caso de los huracanes. Pero en alguna etapa del proceso prebiótico, ya en un escenario de interacciones y transformaciones lo suficientemente rico (es decir, en un marco necesariamente químico), lo novedoso fue que algunos sistemas automantenidos de este tipo pudieron convertirse en sistemas autoproductivos, con un mayor grado de cohesión interna y robustez dinámica, al conseguir generar múltiples constricciones mutuamente dependientes y éstas, a su vez, otras, haciéndose cada vez más complejas y diversas. La base de esta complejización reside en que las constricciones generadas dentro del sistema abren de facto nuevas posibilidades organizativas, al modular selectivamente las dinámicas microscópicas que tienen lugar en el seno del sistema.
«La vida no sólo organiza componentes materiales preexistentes, sino que genera y regenera continuamente la mayoría de ellos y, al tiempo, pasa a controlar buena parte de las condiciones en las que dicha organización dinámica tiene lugar»
En este contexto, el automantenimiento puede comenzar a desplegar además una nueva dimensión: la autorreproducción. En realidad, la autorreproducción no es sino una forma especial de autoproducción. Pero la aparición de la vida requiere que los sistemas automantenidos adopten esa forma de autoproducción que genera nuevas unidades, ya que sólo generando suficiente proliferación espacial y una continuidad temporal indefinida (es decir, una historia abierta) basada en el encadenamiento causal entre organizaciones individuales efímeras se puede asegurar la fijación y el eventual incremento de las innovaciones que fueran apareciendo por azar. Lo que retiene estas innovaciones es simplemente el hecho de que contribuyen al mantenimiento de las organizaciones individuales en las que surgen. Por tanto, su funcionalidad y potencial ventaja selectiva nunca podrán explicarse plenamente fuera del marco de esa organización. Y es esto, junto con el hecho de que haya proliferación y transmisión hereditaria, lo que propicia el comienzo de una forma primitiva de evolución por selección natural.
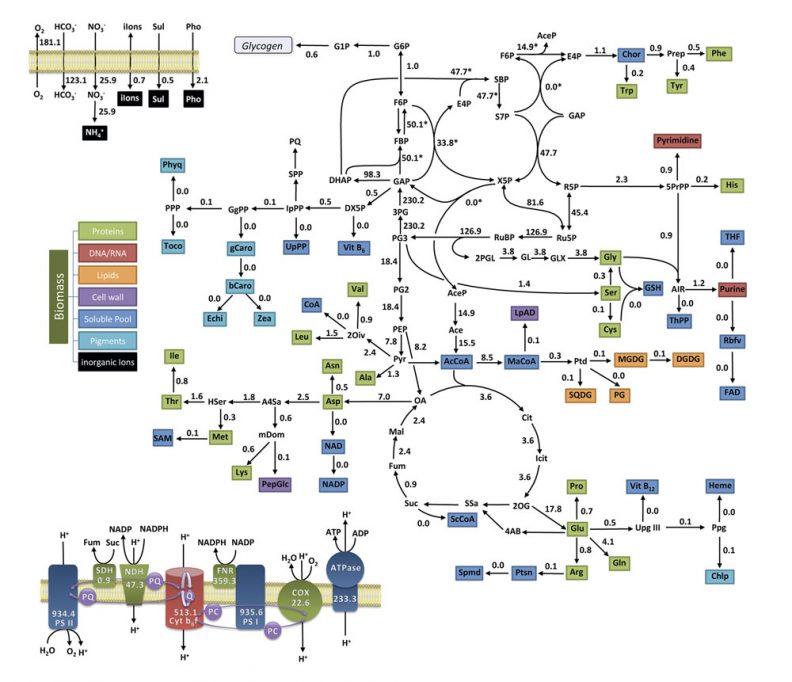
En resumen, no hay «magia» –como pretende Nagel– en el origen de la vida. No hay magia, pero sí estamos ante un fenómeno extraordinariamente intrincado y con profundas implicaciones, tanto científicas como filosóficas, puesto que resulta, en un sentido importante, inexplicable desde la lógica «habitual» que los físicos y químicos usan para entender los fenómenos no vivientes. La emergencia de sistemas materiales capaces de automantenerse de forma robusta y de llevar a cabo ciclos reproductivos estables (es decir, metabolismos genéticamente instruidos) lleva consigo el establecimiento de complejos bucles de transformación química (véase figura 4) que permitan la integración de mecanismos de regulación (basados en relaciones de control jerárquico entre diversos módulos o subsistemas funcionales) junto a mecanismos de herencia fiable (basados en relaciones informacionales). Una teoría general y consistente sobre la organización biológica, en caso de que sea factible elaborarla, debería explicar cómo pueden surgir sistemas naturales que generen y mantengan este tipo de arquitecturas de control regulativo e informacional, tomando en cuenta para ello nuevos resultados provenientes de la química/biología de sistemas y la biología sintética.
«No hay magia en el origen de la vida, pero sí estamos ante un fenómeno extraordinariamente intrincado y con profundas implicaciones, tanto científicas como filosóficas»
Aquí, sin pretender adelantarnos a dichos resultados, hemos tratado de identificar alguna de las problemáticas conceptuales fundamentales que intuimos que surgirán en ese empeño y el tipo de planteamiento teórico que se requerirá para superarlas. La investigación sobre el origen de vida, en la medida en que trata de conectar el mundo de la física y la química con el de la biología, y en la medida en que está obligada a comprender la emergencia e interrelación de ambas dimensiones del fenómeno (tanto su organización individual –es decir, la aparición de metabolismos celulares– como sus capacidades evolutivas a nivel colectivo –el proceso histórico de diversificación e incremento de complejidad), constituye un campo ineludible para hacer posible, si cabe, dicha teoría en un futuro. Y en términos más amplios, por supuesto, también para comprender las relaciones entre las distintas ramas del conocimiento científico ya establecido. Es decir, hasta que no resolvamos la cuestión del origen no seremos capaces de caracterizar con precisión la independencia (o las formas de interdependencia) de la biología con respecto de la física y la química.




