


Does the language faculty lie in our genes? Traditionally, language was considered as one of the most representative examples of cultural transmission (as opposed to biological transmission), a purely cultural phenomenon. From the second half of the twentieth century, though, that conception was dramatically questioned, by asserting that the language faculty is indeed biologically rooted. The objective of this article is threefold: first, it provides a brief picture of the arguments, mainly arising from linguistic development, which point to such a biological character. Further, it is argued that the difference between genotype and phenotype also applies to the linguistic field. Finally, evidence which crucially shows the genetic roots of language is reviewed and discussed: the recent discovery of an autosomal dominant gene (named “FOXP2”), whose disruption triggers severe language disorders.
Durante largo tiempo, se estableció una tajante contraposición entre naturaleza (nature) y crianza (nurture); mientras el ser humano era el (único) representante del segundo dominio, las demás especies quedaban confinadas en el ámbito natural. Según tal visión, nuestra especie, frente al resto, habría trascendido los factores genéticos; así, la primacía de los genes se equiparaba con un veto de cualquier tipo de aprendizaje, mientras que liberarse de tal primacía otorgaba una capacidad ilimitada de aprendizaje; precisamente, la imagen tradicional del ser humano (fuertemente antropocéntrica) era el mejor ejemplo de lo que se puede obtener cuando una especie está liberada del control de los instintos. Instinto, pues, se oponía a aprendizaje, existiendo una brecha insalvable entre los dos planos.
El ejemplo paradigmático de la primacía de la crianza en nuestra especie fue el lenguaje. Tal visión, que encontró especial eco en la tradición estructural-antropológica, es caracterizada por Baker (1996): “Esta tradición considera el lenguaje como un fenómeno cultural; por tanto, las lenguas difieren por las mismas razones por las que difieren las culturas. En concreto, pueden ser moldeadas en vías importantes por la visión del mundo, los modelos de interacción y la historia particular de un grupo concreto de personas. Dado que esos factores culturales pueden ser muy diferentes, en la misma medida las lenguas serán muy diferentes”. En suma, el lenguaje se erigía en el principal estandarte de los progresos que el ser humano había logrado mediante la cultura.
Tal concepción respondía a una motivación global muy clara (aún persistente): la resistencia a aceptar una genética del comportamiento; frente al papel diáfano de los genes en el plano fisiológico, la idea de que también son relevantes en la conducta es polémica, porque suele identificarse con el determinismo.
Sin embargo, tal concepción comenzó a cuestionarse en la segunda mitad del siglo XX en numerosos dominios, lingüística incluida, de modo que se fue rechazando el carácter enfrentado de naturaleza y crianza, favoreciendo en su lugar una integración de los efectos de ambas. El biólogo Michael Ruse (1986) señala que “desde una perspectiva darwinista general, suponer que una cultura está aislada en la cima de los genes es ineficiente y peligroso para ambas partes. La mente como tabula rasa requiere un cerebro con una gran capacidad en blanco […]. Esta receptividad total requeriría probablemente una capacidad craneal mucho mayor de la que ahora poseemos. Este requisito convierte a la hipótesis en altamente improbable”. Por tanto, la posesión, adquirida evolutivamente, de diferentes capacidades de aprendizaje aumenta la eficacia biológica (fitness) de los organismos, ya que el aprendizaje es el medio del que disponen los genes para ajustarse a las exigencias cambiantes del entorno. Wilson (1978) defiende claramente la complementariedad de lo innato y lo aprendido: “El potencial de aprendizaje de cada especie parece estar totalmente programado por la estructura de su cerebro, la secuencia de las descargas de sus hormonas y, finalmente, por sus genes. Cada especie animal está “preparada” para aprender ciertos estímulos, impedida de aprender otros y neutral con respecto a otros más”.
Las dos ópticas señaladas, tradicional y moderna, responden a dos concepciones diferentes sobre el aprendizaje: aprendizaje por instrucción y por selección. Según la primera, correspondiente a la óptica tradicional, el mecanismo de aprendizaje consiste en traspasar información desde el exterior (entorno) al interior del organismo, de modo que éste carece de toda capacidad previa, siendo construido de manera paulatina por la experiencia, según interioriza los estímulos pertinentes. Frente a ella, el aprendizaje por selección implica que el organismo desarrolla rasgos compatibles con las capacidades preexistentes del programa genético. La diferencia entre ambas nociones es acusada: mientras el aprendizaje por instrucción descansa únicamente en el papel de la experiencia, encargada de configurar por completo al ser, el aprendizaje por selección relativiza tal papel; sin negar obviamente su importancia, la función de la experiencia es disparar o activar rasgos innatos específicos de dominio que preexisten a ella.
La sustitución de un paradigma por otro permite insertar al ser humano en una posición realista, acorde con el resto de especies (López García, 2002, que remite el proceso comunicativo al propio nivel celular): desde la óptica tradicional, la especie humana era una suerte de isla en mitad del reino animal; isla puramente cultural, carente de cualquier instinto, ubicada sin embargo en el seno de una naturaleza llena de especies con instintos específicos. Como por arte de magia, nos habríamos librado del predominio de los instintos, accediendo así a un aprendizaje ilimitado.
También el lenguaje participa de tal cambio de perspectiva. La idea de que el lenguaje implicaba un fenómeno cultural por excelencia dejaba sin explicar por qué sólo nuestro sistema, frente al resto de la comunicación animal, estaba libre de factores genéticos. De nuevo nos topamos con ese carácter de isla en medio del mar de la naturaleza que nos otorgamos durante largo tiempo. Hoy en día, sin embargo, hay numerosas evidencias sobre la existencia de un aprendizaje “preparado”, para el lenguaje; en suma, sobre la existencia de factores innatos implicados en su adquisición (para un breve repaso ver Lorenzo y Longa, 2003).
Indicios de la predeterminación biológica del lenguaje
Los argumentos manejados (especialmente, por el innatismo de Noam Chomsky) sobre tal predeterminación biológica, se derivan básicamente del análisis de la adquisición del lenguaje en el niño. Con ello, el estudio del lenguaje está en paralelo con el de otros dominios cognitivos humanos, también centrados en el desarrollo como fuente central de información. Tales argumentos dibujan el siguiente panorama.
«Todos los grupos y sociedades humanas poseen lenguaje y éste es exclusivo de nuestra especie. Además, todas las lenguas, plasmaciones de la facultad del lenguaje, se han revelado como sistemas de idéntica complejidad, con independencia del grado de desarrollo o de aislamiento de una sociedad»
Todos los grupos y sociedades humanas poseen lenguaje y éste es exclusivo de nuestra especie. Además, todas las lenguas, plasmaciones de la facultad del lenguaje, se han revelado como sistemas de idéntica complejidad, con independencia del grado de desarrollo o de aislamiento de una sociedad. Por otro lado, existen correlatos neurales estrechamente ligados a esa capacidad. Auditivamente, estamos preparados para discriminar sonidos lingüísticos con mucha mayor eficiencia que los no lingüísticos, en una proporción de tres a uno. El lenguaje está sometido a un período crítico, pudiendo adquirirse únicamente hasta una determinada edad; más allá de ella, la exposición a la experiencia sólo capacita para adquirir un protolenguaje, carente de las propiedades centrales del lenguaje. Hay una gran diferencia entre la adquisición de la lengua nativa y la de segundas lenguas: mientras todos llegamos (patologías aparte) a dominar perfectamente la nativa, independientemente de nivel de inteligencia, educativo, etc., la adquisición de una segunda lengua, superado el período crítico, debe hacerse con esfuerzo y produce gran variación en los resultados. El niño adquiere en edad muy temprana un sistema de gran complejidad como el lenguaje (a los tres años ya posee un gran dominio) en el tiempo en que es incapaz de afrontar tareas cognitivas mucho más simples. El lenguaje y el resto de la cognición son entidades disociables: una deficiencia cognitiva puede acompañar a una función lingüística no afectada y viceversa. Los niños atraviesan por etapas y ritmos que muestran gran concordancia con independencia de la lengua adquirida (también en lenguas de signos) y la “cantidad” de la experiencia no es relevante: algunas culturas, como la nuestra, tienen un estilo de habla dedicado al niño (maternés), mientras que otras ignoran al niño hasta que puede hablar con solvencia; a pesar de ello, en ambos casos adquieren el lenguaje sin diferencias temporales. El niño puede desarrollar sistemas gramaticales complejos a partir de una evidencia pobre, oral o gestual. Los enunciados lingüísticos, recibidos linealmente, ocultan la estructura formal de las oraciones, que es jerárquica; a pesar de ello, el niño la aprende muy rápido sin ningún tipo de instrucción, desarrollando un sistema intrincado y falto de correspondencia con las señales a las que está sometido (pobreza del estímulo).
«Todos estos hechos, tomados en conjunto, hacen inviable sostener que aprender a hablar implica una mera interiorización de hábitos surgidos en el marco de lo cultural, apuntando a que el lenguaje es un rasgo biológicamente asentado»
Todos estos hechos, tomados en conjunto, hacen inviable sostener que aprender a hablar implica una mera interiorización de hábitos surgidos en el marco de lo cultural, apuntando a que el lenguaje es un rasgo biológicamente asentado.
Genotipo y fenotipo
Si el lenguaje es una facultad con un soporte biológico más que un rasgo cultural (aunque obviamente lo cultural sea relevante para adquirir una lengua concreta), apliquemos la lógica que subyace al aprendizaje por selección. En él, se deben diferenciar dos componentes (cuyos efectos se suman): el estado inicial y la experiencia, que activa al primero. La interacción de ambos factores resulta en un estado estable dado (el rasgo en cuestión). Extendiendo tal visión al lenguaje, el estado inicial es único para la especie, invariable entre los individuos con independencia de la lengua que adquieran (obviamente, es inviable postular diferentes asientos genéticos para diferentes lenguas). Ese estado inicial común a la especie, sometido a una experiencia cambiante (la exposición a una u otra lengua) determina el estado estable (el conocimiento del turco, del gallego o del hindi). La misión del estado inicial es configurar la forma de una lengua humana posible, los rasgos abstractos de diseño de cualquier plasmación del lenguaje, actuando así como guía del proceso de adquisición, que motiva que el niño explore ciertas opciones, pero que descarte automáticamente otras sin considerarlas; de ahí se deriva la rapidez de adquisición, junto al resto de características señaladas.
El genotipo, conformado por el material genético, determina el rango potencial de adaptaciones funcionales al entorno que un organismo puede desarrollar según entre en contacto con diferentes tipos de ambientes. Por su parte, el fenotipo son las propiedades morfológico-estructurales y funcionales desarrolladas en función de la interacción entre los genes del organismo y su entorno. Así, el genotipo, programa interno de desarrollo, se plasma en un fenotipo específico según determinaciones ambientales específicas. Es factible trasladar ambas nociones al lenguaje: los rasgos señalados sobre la adquisición apuntan a un genotipo o estado inicial común, disparado por la experiencia; ésta no llena un organismo “desnudo”, sino que activa respuestas internas al organismo. Tomando un paralelismo, la formación del eje de la estructura del Fucus sp. (un tipo de alga) necesita un estímulo externo, como la luz, pero ese estímulo no es el responsable de tal formación, que depende de la organización interna. Por tanto, la luz es un disparador que pone en marcha algo que está preparado para salir.
«En el estado inicial lingüístico no están especificadas las propiedades de las diferentes lenguas, siendo el papel de tal estado descartar de antemano la posibilidad de que se desarrollen propiedades formales concebibles desde premisas lógicas, pero incompatibles con él»
El estado inicial común a todas las lenguas, la dote genética del lenguaje, no se halla a buen seguro directamente codificada en el genoma, aunque sí determinada por su composición. Monod (1970) señaló que para algunos era una contradicción afirmar que el genoma define por entero la función de una proteína y al tiempo que el contenido informativo de tal función sea mucho más rico que el propio contenido del genoma. Sin embargo, el enriquecimiento informativo, desde la codificación genética hasta su expresión final, implica realizar una estructura somática particular entre otras muchas concebibles a priori, pero irrealizables a partir de tales condiciones iniciales. Por ello, la información final no tiene por qué estar especificada en el genoma, que no obstante descarta otras opciones de desarrollo. De modo similar, en el estado inicial lingüístico no están especificadas las propiedades de las diferentes lenguas, siendo el papel de tal estado descartar de antemano la posibilidad de que se desarrollen propiedades formales concebibles desde premisas lógicas, pero incompatibles con él. Los diferentes estados finales obtenidos o lenguas son sistemas diferenciados entre sí, pero sujetos a un diseño o esquematismo común, constituyendo estados alternativos de desarrollo.
Tal panorama se desprende de los datos sobre la adquisición, que apuntan a una clara base innata. Sin embargo, no debemos equiparar innato con estrictamente genético: al lado están los procesos de epigénesis, interacción entre genes y entorno, que suponen procesos de emergencia no bien conocidos. De hecho, es la epigénesis, especialmente rica en el ser humano, la que motiva la ausencia de una relación cerrada entre genes y comportamiento, dando un enorme margen de maniobra al organismo.
Las bases genéticas del lenguaje
Si la facultad del lenguaje tiene sustento innato, debe transmitirse por herencia. Como es bien sabido, los genes no pueden codificar directamente principios funcionales (tal es, de hecho, la principal reticencia sobre la genética del comportamiento); su misión es codificar la síntesis de las proteínas. Jackendoff (2002) señala que tales proteínas establecen parámetros de desarrollo cerebral aún desconocidos; por ello, por ahora no hay esperanza de comprender los mecanismos indirectos operantes en la transmisión genética del lenguaje (tampoco la del resto de niveles de organización funcional).
| ¿Por qué el alelo mutado es autosómico y dominante?
Gopnik afirmó que el gen implicado en el trastorno específico del lenguaje es autosómico (ligado a uno de los 22 pares de cromosomas no sexuales) y dominante. Gopnik (1990) estudió tres generaciones de una familia inglesa, denominada KE, algunos de cuyos miembros (16 de 31) mostraban problemas con el lenguaje de carácter específicamente gramatical, consistentes en que no disponían de reglas generales sobre rasgos gramaticales (plural, tiempo pasado, género, etc.). El trastorno afectaba aproximadamente al 50% de los miembros de la familia KE (16 de 31). Tal porcentaje imposibilita que el alelo en cuestión sea de naturaleza autosómica recesiva; en este hipotético caso, la probabilidad de transmitir el gen patógeno de cada progenitor es del 50%, y por ello, la de recibirlo de los dos progenitores y manifestar los efectos sería del 25%, no del 50%, como sucede en la familia. Tampoco se puede considerar recesivo, pero ligado al cromosoma X, porque en este caso, el varón podría manifestar sus efectos con una probabilidad del 50% con que sólo la madre fuera portadora; sin embargo, la probabilidad en ese caso para la mujer sigue situada en el 25%. Frente a ello, los casos de mujeres afectadas en la familia son superiores a los de hombres. De las dos posibilidades restantes, hay que descartar su naturaleza dominante pero ligada al cromosoma X, porque este supuesto implicaría que no debería haber mujeres afectadas sólo por transmisión paterna (el gen sano de la madre bastaría para ocultar el efecto); sin embargo, fueron descritos dos de tales casos en la familia (figura 1). Por ello, el gen es de naturaleza autosómica y dominante, lo cual casa con su incidencia en el 50% de la población familiar y con su distribución equitativa entre varones y hembras . |
Pero desde premisas menos ambiciosas, ¿hay alguna evidencia que muestre claramente una base genética para el lenguaje? Dadas las limitaciones en el estudio cerebral, la investigación de esa cuestión, como sucede con otros muchos estudios para entender la localización y funciones de los genes, se centra en localizar deficiencias hereditarias que impliquen fallos en el mecanismo de transmisión del lenguaje. La lógica es clara: “si nuestra facultad lingüística tiene un componente innato, entonces debe haber variación genética para ese rasgo” (Maynard Smith y Szathmáry, 1999). Y para apreciar los efectos, es necesario que el citado menoscabo sea específico del lenguaje, independiente de otros aspectos de la cognición.
Tal evidencia existe y se relaciona con el fenómeno conocido como trastorno específico del lenguaje. Gopnik (1990) estudió tres generaciones de una familia inglesa, denominada KE, algunos de cuyos miembros (16 de 31) mostraban problemas con el lenguaje de carácter específicamente gramatical, consistentes en que no disponían de reglas generales sobre rasgos gramaticales (plural, tiempo pasado, género, etc.). Por ejemplo, producen plurales correctos para palabras conocidas, pero carecen de una regla general de formación de plural capaz de operar con palabras inventadas o no conocidas: no pueden decidir si el plural de una palabra inventada como wug es wugs y deben aprender la forma plural como una pieza léxica diferente en cada caso.
El estudio posterior de familias de otros países reveló que, aunque el trastorno es en parte heterogéneo, existe un núcleo común, conformado por 1) dificultades para adquirir el lenguaje en ausencia de factores susceptibles de ser responsabilizados de ellas (sordera, retraso cognitivo, daño neural aparente o autismo); 2) problemas motores (movimientos orofaciales) y específicamente gramaticales; mientras los primeros son reversibles y se pueden superar con el tiempo, los segundos persisten; y 3) incapacidad para usar las reglas productivas de formación de palabras y para satisfacer todo tipo de requisitos de concordancia. La singularidad del trastorno, pues, estriba en el carácter específicamente gramatical de sus síntomas, así como en su distribución entre los miembros de las familias afectadas (figura 1).
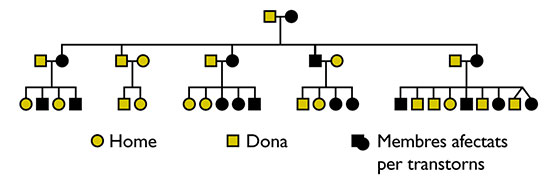
Figura 1. Árbol familiar de tres generaciones de la familia KE. Fuente: Gopnik et al., 1997: 126
Gopnik atribuyó los trastornos lingüísticos de la familia KE a la herencia; en concreto, a un alelo autosómico dominante. Por otro lado, la suposición de que el defecto se relaciona con un solo gen, no con varios, deriva de que el síndrome no se produce en grados.
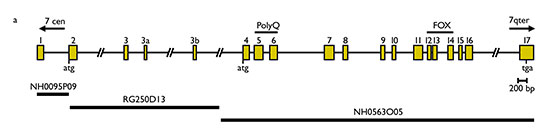
Figura 2. Representación de la estructura del gen FOXP2. La sustitución de guanina por adenina producida en los individuos afectados de la familia KE se localiza en el exón 14. Fuente: Lai et al., 2001: 520
La vinculación del caso descrito con un gen se demostró posteriormente, en dos pasos sucesivos. En 1998, un artículo de un equipo de genetistas (Fisher et al., 1998) expone los resultados de una investigación efectuada en el material genético de esa familia. Detectó una región en el cromosoma 7, SPCH1, que en los miembros afectados presenta una composición diferente a la que rige tanto en los no afectados como en el resto de la población. Este resultado fue concretado aún más por Lai (Lai et al., 2001), que localiza el defecto de la familia KE en un gen de la región SPCH1, bautizado como FOXP2, el primero específicamente vinculado con el lenguaje. Esa localización fue corroborada mediante el examen de otro individuo (CS) sin relación con la familia; por otro lado, análisis genéticos descartaron la coincidencia casual o la posibilidad de un polimorfismo natural: en todos los miembros afectados de la familia (además de en CS), pero en ninguno de los no afectados y en ninguno de los 364 cromosomas analizados de personas no afectadas sin relación con la familia, un nucleótido de guanina aparece reemplazado por uno de adenina, lo que causa un cambio de aminoácidos (la arginina es sustituida por la histidina) que altera la función de la proteína (figuras 2 y 3). Los autores defienden que la alteración en ese gen, de tipo regulador, produce un desarrollo anormal en la embriogénesis de las estructuras neurales relevantes para el lenguaje.

Figura 3. Representación de las tres proteínas conocidas FOXP (1, 2 i 3), junto a las que se reflejan proteínas de otras ramas de la familia FOX. La flecha vertical pequeña (encima de Helix 3) indica el lugar exacto de la sustitución en FOXP2 de la arginina en los miembros afectados de la familia KE. Fuente: Lai et al. 2001: 521
El significado del descubrimiento
Sea cual sea su función exacta, el hallazgo de un gen relacionado con el lenguaje tiene gran trascendencia: por un lado, muestra la factibilidad de una genética del comportamiento, que no implica determinismo, sino aprendizaje “preparado”. Por otro, confirma lo que dicta el sentido común: tanto nuestro sistema comunicativo, como, desde bases más amplias, nuestra especie, no son en cada caso una “isla cultural” situada en medio de un mar de instintos, sino que participamos del resto de especies en tanto que formamos parte de la misma naturaleza. Por ello, el camino abierto mediante el descubrimiento es enormemente prometedor, si bien debe valorarse en sus justos términos. En primer lugar, a buen seguro deben existir otros segmentos genéticos relacionados con el lenguaje, pero no conocidos y quizás no tan fáciles de localizar, teniendo en cuenta fenómenos como la pleitropía o efectos derivados de poligenes. Sin embargo, no es necesario suponer una gran carga genética para el lenguaje, dada la relevancia de los procesos de epigénesis. Por otro lado, debe recordarse que las instrucciones contenidas en los genes son, estrictamente hablando, instrucciones para la especialización celular en tipos particulares de tejidos, no pudiendo plasmar directamente principios funcionales. Por ello, estudios como los citados deben interpretarse como que confirman la existencia de genes que codifican para elaborar tejido neural específicamente dedicado al lenguaje. Los años venideros serán, a buen seguro, apasionantes en este sentido.
BIBLIOGRAFÍA
Baker, M., 1996. The polysynthesis parameter. Oxford University Press. Nueva York.
Fisher, S., Vargha-Khadem, F.,Watkins, K., Monaco, A. y M. Pembrey, 1998. “Localisation of a gene implicated in a severe speech and language disorder”, Nature Genetics, 18.
Gopnik, M., 1990. “Feature-blind grammar and dysphasia”, Nature, 344.
Gopnik, M., Dalalakis, J. y S. Fukuda, 1997. “The biological basis of language: Familial language impairment”. In Gopnik, M. [ed.]. The inheritance and innateness of grammars. Oxford University Press. Nueva York.
Jackendoff, R., 2002. Foundations of language. Brain, meaning, grammar, evolution. Oxford University Press. Nueva York.
Lai, C.S.L., Fisher, S. E., Hurst, J. A., Vargha-Khadem, F. i A. P. Monaco, 2001. “A forkhead domain gene is mutated in a severe speech and language disorder”, Nature, 413.
López García, A., 2002. Fundamentos genéticos del lenguaje. Cátedra. Madrid.
Lorenzo, G. y V. M. Longa, 2003. Homo Loquens. Biología y evolución del lenguaje. TrisTram. Lugo.
Maynard Smith, J. y E. Szathmáry, 1999. The origins of life. From the birth of life to the origin of language. Oxford University Press. Oxford. [Trad. de J. ROS, 2001. Ocho hitos de la evolución. Del origen de la vida a la aparición del lenguaje. Tusquets. Barcelona].
Monod, J., 1970. Le hazard et la nécessité. Du Seuil. París. [1993. El azar y la necesidad. Tusquets, 5a ed. Barcelona].
Ruse, M., 1986. Taking Darwin seriously. Blackwell. Oxford. [1994. Tomándose a Darwin en serio. Implicaciones filosóficas del darwinismo. Salvat. Barcelona].
Wilson, E. O., 1978. On human nature. Harvard University Press. Cambridge, MA. [1980. Sobre la naturaleza humana. FCE. Madrid].




