

Una finestra oberta al funcionament del cervell
Aportaciones de las técnicas de neuroimagen a la neurociencia cognitiva
El ser humano es curioso por naturaleza. Quiere, incluso necesita, saber cómo funciona todo lo que le rodea. Desde niños nos maravillamos cuando, al desmontar un aparato, vemos lo que hay dentro y nos preguntamos para qué sirve tal o cual componente. Y lo que es más asombroso: cuanto más excepcional es la función de un aparato más necesitamos saber de él.
Este símil bien puede aplicarse a nuestro cerebro por su excepcional papel en nuestra existencia y adaptación a un mundo cada vez más complejo. ¿Quién no se ha preguntado alguna vez por alguna de las imponentes funciones cerebrales? ¿Por qué pensamos, por qué tenemos sentimientos tan profundos como el amor, la compasión o la ira, por qué somos creativos, por qué…? Pero lo más importante no es tanto saber el por qué, sino de qué forma el cerebro es capaz de elaborar todas estas capacidades tan complejas del ser humano.
Pues bien, la neurociencia cognitiva es una disciplina relativamente joven que desde la década de los noventa del siglo pasado está respondiendo, al menos parcialmente, a estas preguntas y formulando otras; y, para ello se ha valido de potentes técnicas llamadas, de forma genérica, «técnicas de neuroimagen funcional». La asociación entre neuroimagen y neurociencia cognitiva permite relacionar actividad cerebral y procesos cognitivos siguiendo la siguiente lógica: las áreas cerebrales que muestran mayor activación durante la realización de una tarea cognitiva son cruciales para la función cognitiva que requiere dicha tarea.
«La Neurociencia cognitiva es una disciplina relativamente joven que estudia de qué forma el cerebro es capaz de elaborar todas las capacidades del ser humano»
Así, la conjunción de modelos que intentan estudiar el correlato neural de la cognición y los datos revelados por las técnicas de neuroimagen ha puesto de manifiesto la existencia de una gran cantidad de áreas cerebrales implicadas en cualquier tipo de procesamiento. Pero el verdadero reto de la neurociencia cognitiva es establecer modelos sólidos sobre la cognición humana a partir de los cuales puedan diseñarse experimentos que den respuesta a cómo diferentes áreas cerebrales interconectadas funcionan para producir conducta.
Breve recorrido por las técnicas de neuroimagen
Las técnicas de neuroimagen funcional revolucionaron la neurociencia cognitiva, al permitir estudiar en tiempo real los cambios fisiológicos que ocurren en el cerebro de un individuo mientras realiza una tarea cognitiva (leer oraciones, tomar decisiones, ver imágenes, etc.). En otras palabras, la neurociencia cognitiva combina las estrategias experimentales de la psicología cognitiva con varias técnicas dedicadas a examinar cómo la función cerebral sustenta la actividad mental.
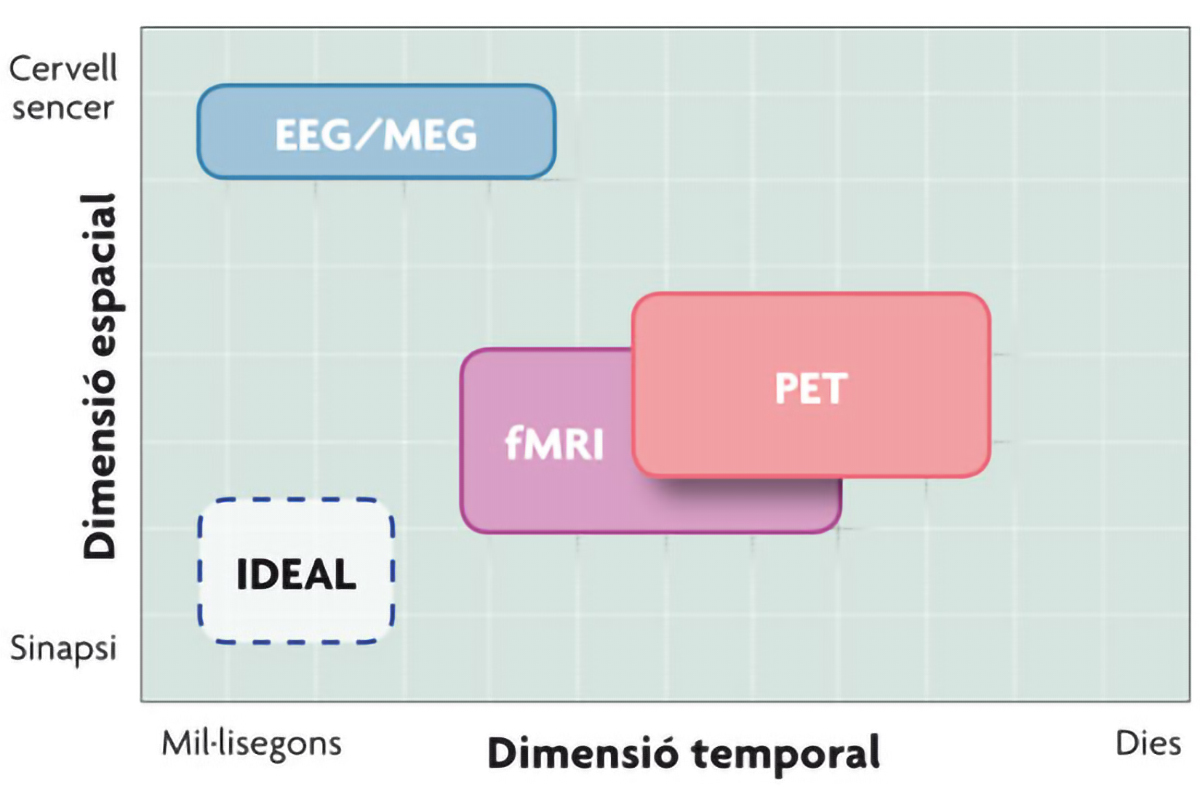
Las técnicas que lideran la investigación actual acerca del funcionamiento cerebral son: los potenciales evento-relacionados (ERP, del inglés Event-Related Potentials), obtenidos a partir del registro electroencefalográfico (EEG), la magnetoencefalografía (MEG), la tomografía por emisión de positrones (PET, siglas de la nomenclatura en inglés, Positron Emission Tomography), y la imagen por resonancia magnética funcional (fMRI, del inglés functional Magnetic Resonance Imaging).
Estas cuatro técnicas permiten registrar la actividad cerebral y se clasifican en dos grandes grupos, en función de la naturaleza de los procesos biológicos que dicha actividad lleva asociada. Dado que el impulso nervioso, la señal fundamental de la comunicación neuronal, se manifiesta a nivel eléctrico, y por tanto también magnético, un primer grupo de técnicas de registro de la actividad cerebral lo constituyen las técnicas magnetoeléctricas: EEG y MEG. Estas representan una medida directa de la actividad neuronal, ya que miden los campos electromagnéticos generados por el flujo de corriente intracelular producido en las neuronas piramidales corticales. Los potenciales evento-relacionados constituyen variaciones del registro magnetoelectroencefalográfico basal, relacionadas con la aparición de un evento y constituyen un indicador neurofisiológico del procesamiento subyacente a esos estímulos.
«El vertader repte de la neurociència cognitiva és establir models sòlids sobre la cognició humana a partir dels quals puguen dissenyar-se experiments que expliquen com diferents àrees cerebrals interconnectades funcionen per a produir conducta»
Por otro lado, las neuronas que se están comunicando activamente necesitan nutrientes para funcionar y, dada su incapacidad para almacenarlos, el cerebro depende del flujo vascular. Así, el incremento regional de la actividad neural se asocia con un incremento local del metabolismo y el riego sanguíneo en esa región cerebral. Las técnicas hemodinámicas, principalmente fMRI y PET, se encargan de medir la actividad de origen metabólico que se produce en determinadas partes del cerebro como consecuencia de la actividad neuronal; es decir, la actividad de un área del cerebro se infiere a partir de la cantidad de glucosa y oxígeno que consume, sustancias que le llegan a través del flujo sanguíneo.
El principio que subyace a ambas técnicas es el mismo, es decir, aquella parte (o partes) del cerebro encargada de llevar a cabo una determinada tarea cognitiva va a aumentar su actividad (y por tanto el consumo de glucosa y oxígeno) con respecto a la actividad de fondo. El hecho de que los procesos que acompañan a estas activaciones cerebrales sean distintos, como acabamos de ver, hace que estas técnicas también difieran en su resolución. Las técnicas que dependen de la hemodinámica cerebral cuentan con una resolución temporal muy limitada, ya que el incremento en el flujo sanguíneo comienza unos segundos después del inicio de la actividad neuronal; pero, por otro lado, el hecho de que no haya una sola neurona en el cerebro a la que no llegue la red de capilares sanguíneos confiere a estas técnicas una alta resolución espacial. En cambio, las técnicas magnetoeléctricas presentan una alta resolución temporal, ya que informan sobre los cambios cerebrales milisegundo a milisegundo, pero su resolución espacial es menor, ya que carecen de capacidad para informar con precisión de las áreas cerebrales activadas.
Una solución para salvar este obstáculo consiste en la combinación de técnicas. Por ejemplo, la combinación de potenciales evento-relacionados y fMRI permite aprovechar la gran resolución temporal de la primera y la gran resolución espacial de la segunda. Este adelanto ha sido posible gracias a la invención de electrodos no ferromagnéticos que pueden ser introducidos en el fuerte campo magnético que se genera dentro del escáner de fMRI.
La importancia del diseño y la interpretación de resultados
Como hemos visto, las técnicas de registro de la actividad cerebral nos proporcionan información acerca de las zonas del cerebro que se activan durante la realización de diversas tareas cognitivas, así como de la temporalidad de los procesos que se llevan a cabo. Sin embargo, el mero conocimiento de la localización de las piezas de un aparato electrónico, por ejemplo, y, el momento en que se pone en marcha, sirve para tener una idea de cómo funciona ese aparato, pero no proporciona una imagen completa de su funcionamiento; es necesario conocer también el programa básico de funcionamiento que gestiona la actividad de los distintos componentes.
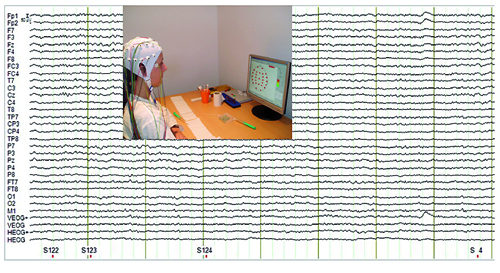
Para realizar un registro electroencefalográfico es necesario colocar unos electrodos en la superficie de la cabeza, que serán los encargados de detectar la actividad cerebral subyacente. Además, se lleva a cabo un registro oculográfico que permite detectar movimientos oculares que pueden contaminar el registro, para eliminarlos posteriormente. Después de conseguir bajar hasta niveles aceptables (alrededor de 3KOhms) la resistencia que ofrecen las diferentes capas que hay desde el tejido cerebral (fuente de la actividad que queremos registrar) a la superficie (lugar donde colocamos los electrodos), comienza el registro de la actividad cerebral ligada a los estímulos presentados./© Pilar Casado y Francisco Muñoz
Esta analogía es válida para el estudio del funcionamiento cerebral. Es evidente que hemos avanzado mucho en el estudio de la función cerebral gracias a los datos que nos ha proporcionado el desarrollo de tecnologías muy avanzadas. Pero no es menos cierto que las interpretaciones e inferencias realizadas a partir de dichos resultados, con el fin de obtener una idea sobre el funcionamiento cognitivo, no siempre han sido acertadas.
Conocido es el «estudio» realizado por Craig Bennet, neurocientífico especializado en el campo del neuromarketing, en el que colocó a un salmón muerto en un escáner y le pidió que se fijara en las imágenes que se mostraban en pantalla y tratara de identificar las emociones en las caras de las personas que aparecían en ellas. Lo curioso es que, por puro azar, aparecieron señales de actividad, justamente en partes del cerebro del salmón; obviamente, no había ninguna actividad en ese cerebro, sino que se produjeron falsos positivos, pequeños errores de medición que a veces afectan a alguno de los vóxels, de lo que se podría haber deducido que un salmón muerto puede identificar emociones humanas. Es evidente que no se pueden extraer conclusiones de este estudio acerca de las «funciones cognitivas» del salmón del Atlántico.
Así, se hace esencial tener presente que los datos obtenidos con las técnicas de neuroimagen han de ser interpretados bajo las premisas de al menos dos principios. En primer lugar, debe existir un modelo teórico sólido y contrastado en el que basar los estudios sobre la función cognitiva, esto es, conocer la actividad cerebral que subyace a una determinada tarea ha de tener como punto de partida un marco teórico que sustente los resultados y que permita integrarlos de manera global. Además, se debe tener en cuenta que en los experimentos de neuroimagen son necesarios una metodología y unos diseños experimentales muy concretos y acotados, que implican el aislamiento de diferentes componentes del procesamiento cognitivo, para abordar mejor su estudio. Pero esto puede conducir a una concepción errónea sobre el supuesto carácter «modular» y «encapsulado» de la cognición.
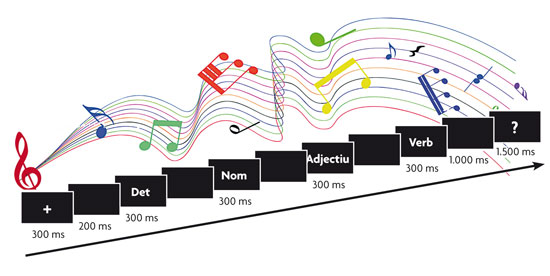
El paradigma experimental en los estudios de procesamiento de oraciones con potenciales relacionado con potenciales evento-relacionados (ERP) suele implicar la presentación de estas oraciones palabra por palabra, una de las cuales presenta una anomalía semántica o sintáctica y constituye el estímulo diana sobre el cual se analiza el potencial relacionado con un acontecimiento. Además, podemos incluir tareas secundarias (escuchar música en este ejemplo), para ver cómo afectan estas al procesamiento./© Pilar Casado y Francisco Muñoz
Siguiendo estas directrices, no son pocos los avances conseguidos en el estudio de las bases neurales de las funciones cognitivas. Por citar solo una de ellas, que a los autores nos resulta particularmente cercana, en los últimos años se han ampliado los modelos de las bases neurales del lenguaje, sustentados básicamente en estudios de lesiones que establecían que el lenguaje es el producto de la actividad de una serie de centros y conexiones entre ellos, que se correspondían esencialmente con el área de Wernicke, el área de Broca y su unión mediante el fascículo arqueado. Así, recientes estudios con fMRI acerca del procesamiento del lenguaje establecen que este se produce siguiendo un flujo continuo de información, que se difunde desde zonas perisilvianas, fundamentalmente en el hemisferio izquierdo, hacia extensas regiones de la corteza cerebral (en realidad, casi todas las regiones corticales parecen susceptibles de participar). Esto incluye por ejemplo la implicación del hemisferio derecho en el procesamiento de aspectos prosódicos del lenguaje, o incluso de zonas frontales motoras en el procesamiento semántico de palabras de acción. Por otra parte, los paradigmas experimentales que permiten el uso de la técnica de los potenciales evento-relacionados (ERP) para el estudio de anomalías lingüísticas, como el que muestra la figura de esta página, han permitido identificar una serie de componentes de los ERP, indicadores de los procesos subyacentes, que están ayudando a esclarecer si los procesamientos semántico y sintáctico interaccionan entre sí, o no, y en ese caso, cuál ocurre primero.
Además, la combinación de nuevas técnicas de imagen cerebral y los modelos más clásicos permiten, sin lugar a dudas, estudiar el procesamiento del lenguaje desde una perspectiva comparada. La tractografía por resonancia magnética es un novedoso método con el que se estudia la orientación de los tractos nerviosos de sustancia blanca <em>in vivo</em>, es decir, sirve para explorar las interconexiones entre diferentes áreas de función cerebral detectadas mediante fMRI. Con esta técnica se ha comprobado que el fascículo arqueado parece haber evolucionado en el cerebro humano a partir de pequeños tractos que conectan las zonas de Broca y Wernicke, ya presentes en el cerebro del macaco, más abundantes en el chimpancé, y con el máximo grado de desarrollo en el humano. En este sentido, una de las principales diferencias entre los cerebros del ser humano y del chimpancé es la notable expansión de las ramificaciones posteriores del fascículo arqueado, particularmente y notablemente dentro el hemisferio izquierdo, lo que al parecer ha desempeñado un papel clave en la evolución del lenguaje humano.
«S’ha posat de manifest l’existència d’una gran quantitat d’àrees cerebrals implicades en qualsevol tipus de processament»
Pero este es solo un ejemplo de cómo se está llevando a cabo esta nueva forma de estudiar el funcionamiento del cerebro; lo que ha conducido a realizar experimentos que nos permiten estudiar aspectos hasta ahora inexplorados, como por ejemplo las emociones y cómo estas influyen en las relaciones sociales. En estudios recientes del grupo del profesor Semir Zeki se ha descrito que existe un único patrón de activación en el cerebro cuando procesamos caras de personas que odiamos, distinto del patrón obtenido cuando procesamos caras de personas que amamos, pero existen áreas comunes. Esto nos da una idea de por qué amor y odio están tan unidos entre sí en nuestra vida diaria. Además, cuando sentimos amor u odio se desactivan ciertas áreas corticales implicadas en el razonamiento y la toma de decisiones, por eso no vemos los estímulos amados u odiados racionalmente.
De todo lo anterior se concluye que, para hacer una correcta interpretación de los resultados en forma de mapas funcionales, deberemos conocer, como estamos argumentando, el modelo teórico que manejamos desde un principio. Con esto queremos decir que la neurociencia cognitiva debe ser consciente de las limitaciones de los modelos y de las técnicas que maneja. En todo caso, se ha de evitar caer en una nueva frenología, intentando asociar funciones cognitivas a un conjunto reducido de áreas cerebrales concretas, ya que cada vez hay más evidencias a favor de que dichas funciones son posibles porque una compleja red de áreas cerebrales se encarga de los procesos neurales que subyacen a tales funciones.
En conclusión, las grandes preguntas de la neurociencia cognitiva acerca de la función cerebral han de estar basadas en la integración de las teorías sobre los procesos que proporciona la psicología cognitiva y la correcta aplicación de las cada vez más potentes técnicas de registro y análisis de la actividad cerebral actualmente disponibles y de las que puedan desarrollarse en el futuro.
Diseño experimental en neuroimagen: un ejemplo con fMRI
La neuroestética constituye una nueva rama del conocimiento neurocientífico que se encarga de estudiar la forma en que el cerebro concibe la belleza, así como la base funcional para crearla, procesarla y entenderla. Esto permite también entender la base neurobiológica de la experiencia artística, la cual constituye, dentro de la inmensa complejidad de las actividades humanas, una más de las que se puede decir que son genuinamente humanas.
Esta disciplina tiene su origen en un estudio pionero (Kawabata y Zeki, 2004) que consistió en estudiar si había áreas específicas del cerebro que se activaban cuando los participantes del mismo visualizaban cuadros de diferentes categorías, retratos, paisajes… que previamente habían considerado como bellos. Los resultados de esta investigación mostraron que la percepción de estas diferentes categorías implicaba la activación de distintas áreas visuales especializadas en el procesamiento y la percepción de cada categoría de estímulo, mientras que, independientemente de la categoría, el hecho de juzgar las pinturas como bellas o no correlacionaba con la activación de estructuras cerebrales específicas, principalmente el córtex orbitofrontal y el córtex motor. De esto se extrae que las percepciones de la belleza y de la fealdad, según este estudio, activan las mismas áreas cerebrales; lo que las diferencia es la intensidad con que se activan.
«La neuroestética constituye una nueva rama del conocimiento neurocientífico que se encarga de estudiar la forma en que el cerebro concibe la belleza»
Sin embargo, la belleza (y la fealdad) no está presente solo en el arte pictórico o de cualquier otro tipo. También puede encontrarse en otros estímulos más naturales y por tanto más adecuados para el estudio de las bases cerebrales del procesamiento estético. Estamos hablando de caras de personas. Siguiendo el planteamiento descrito en el estudio anterior, podemos diseñar un experimento en el que propongamos a nuestros participantes una tarea en la que tengan que clasificar caras de personas, que, previamente (de 3 a 5 días antes de pasar por el escáner), han catalogado como bellas, feas o neutras.
Esto nos permitirá generar una serie de hipótesis en términos de áreas cerebrales implicadas en el procesamiento perceptivo visual, toma de decisiones, y la influencia de las emociones en los procesos cognitivos relacionados con un juicio estético. Si la belleza es un atributo único para nuestro cerebro, el patrón de activaciones encontrado en los dos estudios ha de ser el mismo. Por el contrario, si es dependiente del estímulo, encontraremos un patrón de activaciones específico para las caras.
Con el fin de aceptar (o falsar) esta hipótesis, podríamos diseñar un experimento en el que el participante se introduciría en el escáner de resonancia magnética funcional y los estímulos le serían presentados por unas gafas especiales, a la vez que se le pediría que pulse uno de tres botones, para cada uno de los tres juicios posibles: uno para «bello», otro para «feo» y otro para «neutro». Es muy importante que el diseño experimental cuente con una condición de control formada por estímulos que únicamente varíen en la condición que se pretende estudiar: el carácter estético. El contraste (la diferencia) entre las activaciones cerebrales durante el procesamiento de ambos tipos de conjuntos estimulares dará lugar a la activación neta cerebral, o actividad pura relacionada con los procesos cognitivos y emocionales que estamos estudiando.
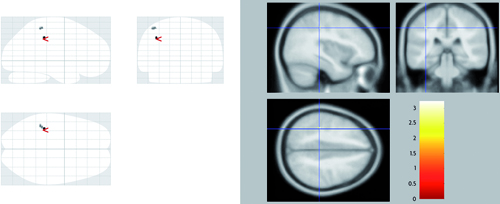
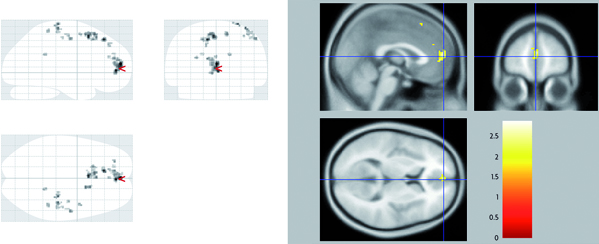
Mapa de activaciones cerebrales esperable durante el experimento. En la figura de la izquierda aparecen todas las áreas que muestran actividad y en las de la derecha, aquellas áreas que han pasado el umbral de significación. En rojo se muestran las áreas más activas durante la percepción de caras feas («bello>feo»), en la figura de arriba, y caras bellas («feo>bello»), en la figura de abajo.
En este caso, un mapa de activaciones esperable sería el que ilustra el recuadro. En las imágenes podríamos observar un conjunto de colores rojo-amarillo sobre las áreas activas, en relación al tipo de contrastes elegidos según nuestra hipótesis de trabajo. El contraste de condiciones «bello > feo», mostraría en rojo las áreas que han estado más activas durante la percepción de imágenes de caras feas. Lo contrario para el contraste alternativo «feo > bello». Lo importante en este punto es que, independientemente del tipo de contrastes, las áreas mostradas se han obtenido usando técnicas estadísticas (como el Statistical Parametric Mapping y muchos otras), que permiten ajustar un nivel de significación concreto (p < 0,05 o incluso p < 0,001) dependiendo de lo riguroso que el investigador quiera ser al descartar activaciones casuales. Fíjese en las figuras de la parte izquierda en las que aparecen todas las áreas que mostrarían actividad. Nuestras inferencias solo deberán basarse en aquellas que hayan pasado el umbral de significación, es decir, las áreas que aparecen a la derecha de las figuras.
De esta forma podríamos razonar que el principal resultado de este experimento sería que el juicio de imágenes de caras bellas sobre las feas lleva consigo la activación de regiones frontales superiores del hemisferio izquierdo. Por otro lado, el patrón de activación durante el juicio de imágenes de caras feas sobre las bellas se asociaría con activaciones en regiones posteriores como el parietal inferior y la cisura calcarina del hemisferio izquierdo.
Podríamos concluir que el juicio estético de una imagen de una cara lleva consigo la activación diferencial de áreas cerebrales, según su belleza/fealdad: en el juicio de la belleza, las áreas activadas son de naturaleza ejecutiva prefrontales, mientras que para el juicio de la fealdad las áreas activadas son de naturaleza perceptiva visual.
Estos resultados coinciden en gran medida con el estudio de Kawabata y Zeki, por lo que podríamos hipotetizar que los humanos contamos con un sentido abstracto de la belleza/fealdad, independientemente de cuál sea la fuente: una obra musical, pictórica o un estímulo visual, como las caras de este estudio.
Para llegar a estas conclusiones y, sobre todo, darlas por válidas, hemos debido diseñar y ejecutar el experimento de forma rigurosa. De hecho, lo primero condiciona decisivamente lo segundo. Y es que estudiar algo tan escurridizo como son las funciones mentales exige un control experimental exquisito.
BIBLIOGRAfíA
Bandettini, P. A., 2009. «What's New in Neuroimaging Methods?». Annals of the New York Academy of Sciences, 1.156: 260-293.
Bartels, A. y Zeki, S., 2000. «The Neural Basis of Romantic Love». Neuroreport, 11(17): 3.829-3.834.
Bennett, C. M. et. al., 2010. «Neural Correlates of Interspecies Perspective Taking in the Post-mortem Atlantic Salmon: an Argument for Multiple Comparisons Correction». Journal of Serendipitous and Unexpected Results, 1(1):1-5.
Buckner, R. L.y J. M. Logan, 2001. «Functional Neuroimaging Methods: PET and fMRI». In Cabeza, R. y A. Kingstone (eds.). Handbook of Functional Neuroimaging of Cognition. MIT Press. Cambridge.
Carretié, L., 2011. Anatomía de la mente. Emoción, cognición y cerebro. Pirámide. Madrid.
Kutas, M., et. al., 2006. «Psycholinguistics Electrified II: 1994-2005». In Traxler, M. i M. A. Gernsbacher (eds.). Handbook of Psycholinguistics. Elsevier. Nova York.
Martín-Loeches, M. P. y Casado, 2012. «Functional and Structural Magnetic Resonance Imaging of Human Language: A Review». In Bright, P. (ed.). Neuroimaging. Cognitive and Clinical Neuroscience. InTech. Rijeka.
Zeki, S. i J. P. Romaya, 2008. «Neural Correlates of Hate». PLoS One, 3(10): e3556.
Kawabata, H. i S. Zeki, 2004. «Neural Correlates of Beauty». Journal of Neurophysiology, 91: 1.699-1.705.




