

El registro fósil de la inteligencia de los primates
De los primeros primates a los orígenes humanos

Los animales recogen y procesan la información necesaria para sobrevivir y reproducirse. El medio que utilizan para asimilarla es la inteligencia, que a su vez va en función del cerebro, su morfología, tamaño, organización y citoarquitectura. Mientras que la organización interna y la interconectividad celular de los cerebros de los animales fosilizados son invisibles para la paleontología, el tamaño y la morfología superficial de su cerebro a veces sí se conserva, normalmente solo de forma parcial, como molde endocraneal (el molde natural o artificial del interior de la cavidad craneal). El estudio de la evolución de la inteligencia en los primates, tal como se interpreta a partir del registro fósil de moldes endocraneales, se centra principalmente en los linajes que nos pueden dar una información más directa sobre los acontecimientos evolutivos que condujeron al origen de la inteligencia humana.
Palabras clave: evolución de los primates, paleoneurología, paleoantropología, hominoideos, cognición.
Desde su establecimiento, el orden Primates se ha considerado especialmente inteligente en comparación con otros animales. Hoy reconocemos capacidades cognitivas similares a las de los primates en otros animales, pero es innegable que los primates priorizan la cognición y la flexibilidad conductual frente a comportamientos determinados genéticamente.
Orígenes de los Euprimates
Los Plesiadapiformes (de hace 65 a 37 millones de años) son el taxón fósil más antiguo atribuido al orden Primates, aunque sus miembros carecen de la mayoría de los atributos que definen a los Euprimates, primates de aspecto moderno (Bloch et al., 2007). Algunos han sugerido que los Plesiadapiformes están más estrechamente relacionados con los Dermoptera (colugos o lémures voladores) (Beard, 1990; Kay et al., 1990), pero el consenso actual es que se trata de primates primitivos. Este grupo increíblemente diverso persistió más de 25 millones de años y se extendió desde Norteamérica hasta Asia y Europa. Los Plesiadapiformes evolucionaron a partir de mamíferos arbóreos y sus extremidades traseras son prénsiles como las de los primates, pero sus dientes se parecen a los de los primeros mamíferos y de los insectívoros actuales, con puntas y bordes afilados para perforar y partir los exoesqueletos de los insectos (Silcox et al., 2015). Aunque tenían dientes similares, estos eran algo más anchos y con puntas más redondeadas, lo que puede indicar que, además de insectos, su dieta incluía fruta y otros productos vegetales (la savia, por ejemplo). Es una diferencia sutil, pero es posible que encauzara a los primates hacia una dieta y una forma de vida más diversas y adaptables.

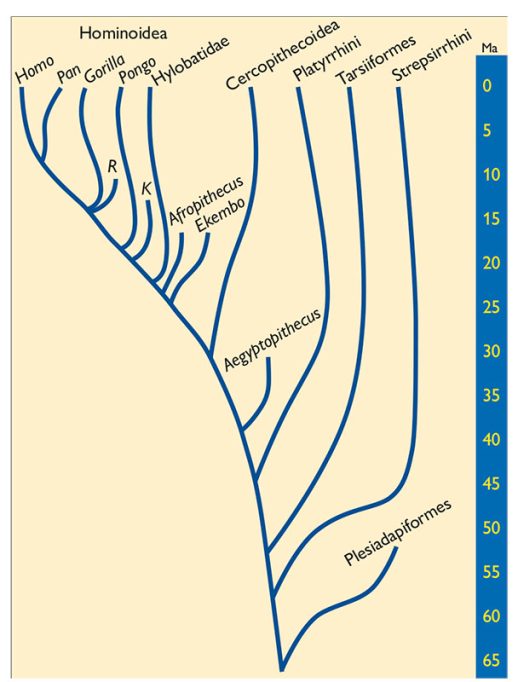
Taxonomía de los primates. Resumen simplificado de la sistemática de los primates. / Fuente: Elaboración propia
El primer fósil descrito de un euprimate (de ahora en adelante, primates), Adapis parisiensis, no se reconoció al principio como primate, sino como artiodáctilo (ungulados de dedos pares como las vacas, las ovejas o los antílopes). Los adápidos, la familia a la que pertenecía Adapis, tenían extremidades flexibles y diestras, y manos y pies prénsiles con uñas y almohadillas grandes, en lugar de garras. Estos primates de aspecto moderno aparecieron en el Eoceno (hace 56–33,9 millones de años) (Godinot, 2015). Al principio, cuando se reconocieron los fósiles de primates como tales, se observó que, al igual que los primates modernos, solían tener cerebros relativamente grandes, hocicos pequeños y ojos grandes y convergentes. Estos rasgos sugieren una menor dependencia del olfato y mayor dependencia de la visión en comparación con otros mamíferos, así como una mayor capacidad cognitiva.
Para tratar de explicar el énfasis táctil y visual de los sistemas sensoriales de los primates, las teorías se han centrado en la dieta y la ecología (Cartmill, 1992; Sussman, 1991). Al igual que los primates modernos, los primitivos pasaban la mayor parte del tiempo en los árboles. Un hábitat arbóreo plantea muchos retos: la tridimensionalidad, la importancia de evitar caídas al saltar y moverse con velocidad entre las ramas y la capacidad de cazar o recolectar alimentos en las ramas más inestables. La caza, en particular la capacidad de atrapar insectos utilizando su visión tridimensional, es posible gracias a la superposición de los campos visuales. Los primeros primates evolucionaron en un contexto de abundancia de vegetación angiosperma (plantas con flores), que no solo producen frutos en las ramas, sino que también atraen a los insectos. Los primeros primates también eran nocturnos –lo sabemos por el gran tamaño de las órbitas oculares–, lo que plantea incluso más retos para cazar y moverse entre las ramas (Wu et al., 2022).
Estos primates ya daban muestras potenciales de una inteligencia aumentada en el tamaño y la morfología del cerebro. En comparación con los mamíferos contemporáneos, los cerebros de los primeros primates eran más grandes en términos absolutos y tenían lóbulos olfativos más pequeños, en relación con el tamaño total del cerebro (Radinsky, 1979). En general, el tamaño del cerebro se considera un indicador indirecto de inteligencia, aunque uno imperfecto (Gibson et al., 2001; Street et al., 2017). La consecución de grados más modernos de primates
–de estrepsirrinos a haplorrinos, de antropoides basales a hominoideos, de hominoideos basales a simios modernos, de simios a humanos– suele estar acompañada de una reorganización de regiones cerebrales, que también se produce cuando se desarrollan mayores niveles de inteligencia (Gibson et al., 2001; Radinsky, 1979; Street et al., 2017).
Entre los primates primitivos con encéfalos bien conservados se encuentran los adápidos y los omomíidos, que se parecen a los lémures y lorinos actuales, y los tarsiformes primitivos, antepasados de los tarseros actuales. Los cráneos más antiguos tenían cerebros más pequeños que los de los prosimios actuales (en relación con el tamaño corporal) pero, hacia el final de su rango geológico, los primeros primates habían alcanzado un tamaño cerebral similar a los más pequeños de entre los prosimios actuales (Godinot, 2015). El aumento del tamaño del cerebro a lo largo del tiempo es común en los linajes de mamíferos y puede explicarse por lo que se conoce como una carrera armamentística, en la que la selección mejora la inteligencia de los depredadores para cazar a sus presas y la de las presas para detectar mejor la presencia de sus depredadores (Ward et al., 2004). El tamaño del cerebro comienza a aumentar de forma más clara con la evolución de los primates antropoides (monos, simios y humanos). Esto va acompañado de un cambio hacia la actividad diurna y la aparición de grupos sociales (Godinot, 2015). La actividad diurna se deduce del tamaño de las órbitas (más pequeñas que en las especies nocturnas), mientras que la sociabilidad se deduce del dimorfismo corporal y del tamaño de los caninos. Sin embargo, por muy importante que la inteligencia social pueda ser para los antropoides actuales, especialmente para los catarrinos (monos del viejo mundo, simios y humanos), la complejidad social no parece ser un predictor tan fiable del tamaño del cerebro como la dieta y la socioecología (López-Aguirre et al., 2022).

Antropoides
Los antropoides, un suborden de primates, comprenden los monos del viejo y el nuevo mundo, los simios, los humanos y sus parientes fósiles. La mayoría de los estrepsirrinos fósiles ya se habían extinguido cuando los antropoides empezaban a ser comunes en el registro fósil (los primeros antropoides son muy poco comunes antes de hace unos 40 millones de años). Las mejores muestras de antropoides primitivos proceden de los yacimientos de El Fayum, en Egipto. Se conocen varios géneros de antropoides primitivos con cráneos relativamente bien conservados (Godinot, 2015).
Los antropoides de El Fayum incluyen una serie de haplorrinos y catarrinos primitivos (Godinot, 2015). Eran arbóreos, algunos principalmente frugívoros y otros más insectívoros. Parece que la mayoría eran diurnos y presentaban dimorfismo sexual en el tamaño corporal y en los caninos, lo que sugiere una complejidad social similar a la de los antropoides actuales. En general, son más grandes que la mayoría de los prosimios del Eoceno. Disponemos de moldes endocraneales de los antropoides primitivos Proteopithecus y Parapithecus y de los catarrinos primitivos Catopithecus y Aegyptopithecus (Godinot, 2015). Los taxones más primitivos tienen cerebros pequeños y lóbulos olfativos grandes, mientras que Aegyptopithecus, más joven y con una morfología craneal más moderna, tiene lóbulos olfativos reducidos y un cerebro comparable a los más pequeños que podemos encontrar en los prosimios modernos. En resumen, los primeros antropoides conservan el patrón primate (antiguo y actual) de arborismo y dieta frugívora, y muestran un cambio hacia un comportamiento más diurno y social, con sutiles cambios en el tamaño y la morfología cerebral que pueden sugerir algunos incrementos en inteligencia.

Cronología de la radiación de los simios en el Mioceno y atributos asociados a su diversidad y evolución. Los taxones se enumeran por orden cronológico dentro de cada subépoca. La tabla omite muchos taxones no tratados en el texto, en particular los grandes simios asiáticos, cuyos cráneos no se conocen bien. Los atributos exclusivos de los grandes simios y los humanos, incluidos los relacionados con la locomoción y el cerebro, evolucionaron en el Mioceno. / Fuente: Elaboración propia
Hominoideos
La superfamilia Hominoidea, parte de los Anthropoidea, incluye a simios, humanos y sus parientes fósiles. Aunque no es el hominoideo más antiguo, nuestro conocimiento de la evolución de los simios comienza realmente con Ekembo, un hominoideo primitivo encontrado en diversas localidades de Kenia, cuya edad oscila entre 19 y 17 millones de años (McNulty et al., 2015). Ekembo es el primer catarrino completamente moderno. Se conocen la mayoría de los huesos de su esqueleto, que revelan una estructura corporal muy parecida a la de los monos actuales, con una excepción importante: un coxis en lugar de cola. La ausencia de cola es un carácter derivado que se considera un sello distintivo de los Hominoidea, y posiblemente los obligó a desarrollar una mayor destreza manual para compensar la ausencia de la cola como mecanismo de equilibrio en los árboles. El cráneo de Ekembo presenta un tamaño cerebral medio dentro del rango de los monos y gibones actuales (Begun y Kordos, 2004). En general, Ekembo se asemeja a un mono en su adaptación global, con atributos simiescos sutiles pero inequívocos. Probablemente era al menos tan inteligente como los monos del viejo mundo actuales y tenía una flexibilidad similar en la búsqueda de alimento, el desplazamiento y los comportamientos sociales.

Afropithecus se solapa temporalmente con la última etapa de Ekembo (Leakey y Walker, 1997). Postcranealmente, ambas especies son indistinguibles. Desde el punto de vista craneodental, Afropithecus se distingue por tener un gran rostro, asociado a grandes músculos masticadores que evidencian mandíbulas y dientes poderosos desarrollados para triturar y moler. Afropithecus también tiene especializaciones en sus dientes frontales que probablemente le permitían desgarrar cualquier cubierta protectora. Es probable que Afropithecus pudiera explotar una mayor variedad de recursos que Ekembo, lo que podría haber permitido a sus descendientes dispersarse hacia el norte, a Arabia Saudí y luego a Europa (Begun, 2015a; 2015b). Fue en Eurasia donde evolucionaron los homínidos modernos (grandes simios y humanos).
Los hominoideos europeos más antiguos son homínidos (orangutanes, chimpancés, bonobos, gorilas, humanos y sus parientes fósiles), con dientes gruesos y esmaltados de aspecto moderno, amplios y planos. Al igual que Afropithecus, los simios europeos como Griphopithecus tenían dietas amplias y podían explotar recursos que iban desde frutos blandos hasta alimentos más complicados con cubiertas exteriores, como cáscaras o piel dura, así como alimentos subterráneos como raíces. La variedad de la dieta fue probablemente la clave del éxito de los primeros homínidos que se dispersaron por Eurasia. Esta dieta era posible gracias a las adaptaciones masticatorias, y puede que fuera acompañada de cambios en la cognición relacionados con la flexibilidad ecológica y dietética. Una vez en Eurasia, los homínidos del Mioceno medio se dispersaron ampliamente, ocupando la región comprendida entre Europa Central y el ecuador. Surgen múltiples taxones con diversas adaptaciones, pero a todos les unen atributos mejorados para el procesamiento de alimentos. Los simios del Mioceno medio que conocemos muestran adaptaciones indicativas de un cambio hacia un comportamiento postural con mayor importancia de las extremidades anteriores, pero sin alcanzar todavía la suspensión ni la bipedestación (véase más adelante) (Nakatsukasa y Kunimatsu 2009). Es posible que realizaran más actividades de escalada vertical y elevación, pero no se suspendían de las ramas como los hominoideos no humanos actuales. No se conocen cráneos de simios del Mioceno medio, pero a finales del Mioceno sí hay pruebas del tamaño de un cerebro y de una biología del desarrollo similar a la de los grandes simios modernos.
Hace unos 12,5 millones de años aparecieron en Europa simios más modernos. El más conocido es Pierolapithecus, encontrado en un yacimiento de 11,9 millones de años de antigüedad cerca de Barcelona (Alba, 2012). Pierolapithecus era bípedo y tenía una columna vertebral más vertical, como los simios actuales –a diferencia de las columnas más horizontales de los monos y de la mayoría de los simios del Mioceno temprano y medio– (Moyà-Solà et al., 2004). Sus manos eran grandes, con dedos largos y curvados, lo que sugiere cierto grado de suspensión. Pierolapithecus y otros simios del Mioceno medio tardío europeo tenían dientes muy parecidos a los de los chimpancés actuales y probablemente seguían una dieta similar, rica en frutos blandos pero con la variedad suficiente para capear los periodos de escasez, explotando otros recursos. Al final del Mioceno medio aparecen otros simios de aspecto moderno, el más famoso de los cuales es Danuvius, del comienzo del Mioceno tardío (Böhme et al., 2019). Danuvius se parece a los simios del final del Mioceno medio, como Pierolapithecus, en la morfología dental. Al igual que este, era bípedo, y tenemos pruebas de sus poderosas manos prénsiles. Las extremidades posteriores de Danuvius indican que era bípedo en los árboles y tenía fuertes pies prénsiles. La bipedestación de Danuvius podría ser precursora de la humana (Böhme et al., 2019).
Sabemos poco sobre el cerebro de estos simios. Conocemos a los simios del Mioceno tardío europeo, como Rudapithecus en Hungría e Hispanopithecus en España, a partir de esqueletos parciales que revelan una estructura corporal moderna, bípeda y totalmente suspensoria. También se conocen tres cráneos parciales de Rudapithecus, y dos de ellos incluyen el neurocráneo. Gracias a ellos sabemos que el cerebro de Rudapithecus estaba dentro del rango del de los chimpancés actuales, si tenemos en cuenta las diferencias de tamaño corporal entre ambos (Begun y Kordos, 2004; Gunz et al., 2020). Dado que el tejido cerebral es uno de los más «caros» en términos metabólicos, es muy probable que el aumento del tamaño del cerebro esté asociado a la selección de múltiples comportamientos y habilidades que mejoran la supervivencia y el éxito reproductivo. Que Rudapithecus hubiera alcanzado este nivel de encefalización, difícil de explicar en ausencia de un beneficio para la supervivencia y la reproducción, sugiere un patrón de comportamiento que se aproxima al nivel de complejidad de los grandes simios modernos. Algunas pruebas indirectas de la evolución de una mayor capacidad cognitiva en los simios proceden del tamaño corporal y, especialmente, del desarrollo dental (Kelley, 2004; Ward et al., 2004). En Sivapithecus (un gran simio del sur de Asia emparentado con los orangutanes) y Rudapithecus, las pruebas dentales indican que ambos taxones se desarrollan a un ritmo comparable al de los grandes simios actuales, que tardan más en alcanzar la madurez que los monos del viejo mundo actuales (Kelley, 2004; Smith et al., 2019). El simio fósil Afropithecus también parece caracterizarse por un crecimiento más lento, aunque no en el grado observado en los taxones del Mioceno tardío (Smith et al., 2003).

Conclusiones
La historia evolutiva de los primates se caracteriza por una diversidad de adaptaciones a un estilo de vida arbóreo, una creciente dependencia de las señales visuales frente a las olfativas y una necesidad cada vez mayor de mejorar las capacidades cognitivas relacionadas con la adquisición y el procesamiento de recursos, las interacciones sociales y la diversidad ecológica. Todos los primates actuales, pero especialmente los antropoides, son excepcionales en sus niveles de inteligencia en comparación con la mayoría de los mamíferos. Diversos logros cognitivos impresionantes caracterizan varias ramas de antropoides. Ejemplo de ello son el uso de herramientas en los monos Cebus y los extraordinarios logros de muchos monos del viejo mundo (lavado de alimentos, baños en arroyos, increíble flexibilidad de comportamiento y capacidad para adaptarse a la presencia de humanos). Los distintos primates han desarrollado sus propias versiones de excepcionalidad cognitiva. Pero el registro fósil de atributos relacionados con la inteligencia muestra su punto álgido en los simios del Mioceno tardío, que alcanzaron la estructura corporal, la biología reproductiva y, probablemente, la cognición de los grandes simios modernos. Estos simios, antepasados en sentido amplio de los homininos, sentaron las bases para la evolución cognitiva de los humanos. El registro fósil de los homininos muestra aumentos constantes del tamaño del cerebro a lo largo del tiempo, correlacionados con el desarrollo de estrategias de supervivencia extrasomáticas progresivamente más complejas y elaboradas; esto es, la cultura humana.
Referencias
Alba, D. M. (2012). Fossil apes from the Vallès-Penedès basin. Evolutionary Anthropology, Issues, News, and Reviews, 21(6), 254–269. https://doi.org/
10.1002/evan.21312
Begun, D. R. (2015a). The real planet of the apes. Princeton University Press.
Begun, D. R. (2015b). Fossil record of Miocene hominoids. En W. Henke & I. Tattersall (Eds.), Handbook of palaeoanthropology (pp. 1262–1332). Springer.
Begun, D. R., & Kordos, L. (2004). Cranial evidence of the evolution of intelligence in fossil apes. En A. E. Russon & D. R. Begun (Eds.), The evolution of thought: Evolutionary origins of great ape intelligence (pp. 260–279). Cambridge University Press.
Beard, K. C. (1990). Gliding behaviour and palaeoecology of the alleged primate family Paromomyidae (Mammalia, Dermaptera). Nature, 345, 340–341. https://doi.org/10.1038/345340a0
Bloch, J. I., Silcox, M. T., Boyer, D. M., & Sargis, E. J. (2007). New Paleocene skeletons and the relationship of plesiadapiforms to crown-clade primates. Proceedings of the National Academy of Sciences, 104(4), 1159–1164. https://doi.org/10.1073/pnas.0610579104
Böhme, M., Spassov, N., Fuss, J., Tröscher, A., Deane, A. S., Prieto, J., Kirscher, U., Lechner, T., & Begun, D. R. (2019). A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature, 575(7783), 489–493. https://doi.org/10.1038/s41586-019-1731-0
Cartmill, M. (1992). New views on primate origins. Evolutionary Anthropology: Issues, News, and Reviews, 1(3), 105–111. https://doi.org/10.1002/evan.1360010308
Gibson, K. R., Rumbaugh, D., & Beran, M. (2001). Bigger is better: Primate brain size in relationship to cognition. En D. Falk & K. R. Gibson (Eds.), Evolutionary anatomy of the primate cerebral cortex (pp. 79–97). Cambridge University Press.
Godinot, M. (2015). Fossil record of the primates from the Paleocene to the Oligocene. En W. Henke & I. Tattersall (Eds.), Handbook of palaeoanthropology (pp. 1137–1259). Springer. https://doi.org/10.1007/978-3-642-27800-6_68-1
Gunz, P., Kozakowski, S., Neubauer, S., Le Cabec, A., Kullmer, O., Benazzi, S., Hublin, J., & Begun, D. R. (2020). Skull reconstruction of the late Miocene ape Rudapithecus hungaricus from Rudabánya, Hungary. Journal of Human Evolution, 138, 102687. https://doi.org/10.1016/j.jhevol.2019.102687
Kay, R. F., Thorington, R. W., & Houde, P. (1990). Eocene plesiadapiform shows affinities with flying lemurs not primates. Nature, 345, 342–344. https://doi.org/10.1038/345342a0
Kelley, J. (2004). Life history and cognitive evolution in the apes. En A. E. Russon & D. R. Begun (Eds.), The evolution of thought: Evolutionary origins of great ape intelligence (pp. 280–297). Cambridge University Press.
Leakey, M., & Walker, A. (1997). Afropithecus: Function and phylogeny. En D. R. Begun, C. V. Ward, & M. D. Rose (Eds.), Function, phylogeny and fossils: Miocene hominoid evolution and adaptations (pp. 225–239). Plenum Press.
López-Aguirre, C., Lang, M. M., & Silcox, M. T. (2022). Diet drove brain and dental morphological coevolution in strepsirrhine primates. PLOS ONE, 17(6), e0269041. https://doi.org/10.1371/journal.pone.0269041
McNulty, K. P., Begun, D. R., Kelley, J., Manthi, F. K., & Mbua, E. N. (2015). A systematic revision of Proconsul with the description of a new genus of early Miocene hominoid. Journal of Human Evolution, 84, 42–61. https://doi.org/10.1016/j.jhevol.2015.03.009
Moyà-Solà, S., Köhler, M., Alba, D. M., Casanovas-Vilar, I., & Galindo, J. (2004). Pierolapithecus catalaunicus, a new middle Miocene great ape from Spain. Science, 306(5700), 1339–1344. https://doi.org/10.1126/science.
1103094
Nakatsukasa, M., & Kunimatsu, Y. (2009). Nacholapithecus and its importance for understanding hominoid evolution. Evolutionary Anthropology: Issues, News, and Reviews, 18(3), 103–119. https://doi.org/10.1002/evan.20208
Radinsky, L. (1979). The fossil record of primate brain evolution. 49th James Arthur Lecture. American Museum of Natural History.
Silcox, M. T., Sargis, E. J., Bloch, J. I., & Boyer, D. M. (2015). Primate origins and supraordinal relationships: Morphological evidence. En W. Henke & I. Tattersall (Eds.), Handbook of palaeoanthropology (pp. 1053–1081). Springer. https://doi.org/10.1007/978-3-642-39979-4_29
Smith, T. M., Martin, L. B., & Leaky, M. G. (2003). Enamel thickness, microstructure and development in Afropithecus turkanensis. Journal of Human Evolution, 44(3), 283–306. https://doi.org/10.1016/s0047-2484(03)00006-x
Smith, T. M., Tafforeau, P., Pouech, J., & Begun, D. R. (2019). Enamel thickness and dental development in Rudapithecus hungaricus. Journal of Human Evolution, 136, 102649. https://doi.org/10.1016/j.jhevol.2019.102649
Street, S. E., Navarrete, A. F., Reader, S. M., & Laland, K. N. (2017). Coevolution of cultural intelligence, extended life history, sociality, and brain size in primates. Proceedings of the National Academy of Sciences, 114(30), 7908–7914. https://doi.org/10.1073/pnas.1620734114
Sussman, R. W. (1991). Primate origins and the evolution of angiosperms. American Journal of Primatology, 23, 209–223. https://doi.org/10.1002/ajp.1350230402
Ward, C. V., Flynn, M., & Begun, D. R. (2004). Body size and intelligence in hominoid evolution. En A. E. Russon & D. R. Begun (Eds.), The evolution of thought: Evolutionary origins of great ape intelligence (pp. 335–349). Cambridge University Press.
Wu, Y., L. Fan, L., Bai, L., Li, Q., Gu, H., Sun, C., Jiang, T., & Feng, J. (2022). Ambush predation and the origin of euprimates. Science Advances, 8(37), eabn6248. https://doi.org/10.1126/sciadv.abn6248




