

Movimientos de dispersión en los primates
Variabilidad en sus patrones y causas

Los movimientos de dispersión en los animales representan decisiones críticas para los individuos, ya que afectan a su supervivencia y éxito reproductivo, además de ser un componente importante de la dinámica poblacional. En este artículo se describe la variabilidad en los patrones de dispersión en los primates y algunas de las causas, tanto últimas como proximales, a las que responde.
Palabras clave: competencia intragrupal, elección de pareja, infanticidio, depresión consanguínea.
«La dispersión proporciona una conexión genética entre las poblaciones e influye de manera importante en los procesos demográficos y en la dinámica poblacional»
La distribución espacial de los organismos no suele ser aleatoria, sino que responde a los factores bióticos y abióticos presentes en el ambiente. Dicha distribución, ya sea agregada o dispersa, es el resultado del movimiento de los individuos hacia la localización que ocupan en cada momento. A este respecto, los conceptos de migración y dispersión se emplean para describir, de manera general, las características de ciertos movimientos que realizan los organismos. La migración hace referencia al movimiento conjunto de un número elevado de individuos de la misma especie entre dos hábitats distintos. Los movimientos migratorios pueden ser cíclicos (con distinta periodicidad, por ejemplo diaria o anual) o de ida únicamente, lo que da lugar a diversos patrones migratorios. La dispersión hace referencia al movimiento que provoca el alejamiento de los individuos de sus áreas natales y/o de sus progenitores e individuos cercanos. Estos movimientos se pueden producir en solitario o en pequeños grupos y pueden ser activos o pasivos. La dispersión es una de las características clave de las poblaciones debido a que proporciona una conexión genética entre estas e influye de manera importante en los procesos demográficos y en la dinámica poblacional. En este artículo nos centraremos en los movimientos de dispersión en los primates.
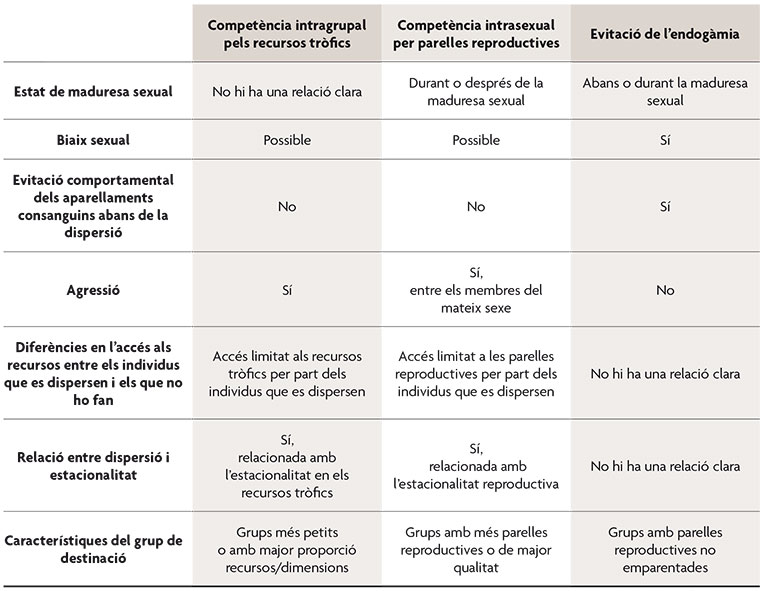
Tabla 1. Características de los patrones de dispersión y su posible relación con las tres principales causas últimas de la dispersión natal. / Adaptado de la Tabla 2 de Field y Guatelli-Steinberg, 2003.
Dispersión en los primates
El orden de los primates, al que pertenece la especie humana, está constituido por especies altamente sociales cuyos individuos viven en grupos, en muchos casos durante toda la vida. A este respecto la dispersión en primates, y en general en las especies sociales, tiene dos facetas; la ecológica (dispersión espacial) y la social (dispersión social) (Isbell y Van Vuren, 1996). En la dispersión espacial los individuos se trasladan a áreas domésticas (home range) desconocidas, mientras que en la dispersión social los individuos se trasladan a nuevos grupos sociales. Estos dos aspectos de la dispersión se pueden producir de manera conjunta o no, dependiendo, principalmente, del tipo de organización social de los grupos. En aquellas especies cuyos grupos defienden un territorio, como en el caso de los chimpancés, la dispersión de los individuos suele combinar ambos aspectos. En otras especies, como los gorilas o los babuinos hamadríades, las áreas domésticas de los grupos sociales se solapan, por lo que, generalmente, la dispersión de los individuos posee un componente social pero no ecológico. Por último, la dispersión espacial en ausencia de dispersión social implicaría el desplazamiento de todo el grupo social a un área doméstica diferente. Estos desplazamientos suelen ser el resultado de alteraciones en el hábitat que afectan a recursos o factores vitales para la especie, lo que impulsa al grupo a buscar nuevos lugares con unas condiciones mejores. Aunque hemos considerado este tipo de movimiento como un caso de dispersión espacial, también podría ser valorado como un proceso migratorio no estacional.
¿Por qué abandonar el entorno natal o conocido?
La incorporación a un nuevo grupo social supone en muchos casos la pérdida de las relaciones sociales cooperativas establecidas en el anterior grupo y la necesidad de establecer nuevas relaciones, con el consiguiente gasto en tiempo y en energía. Todo ello tiene lugar en un entorno social generalmente hostil en el que los miembros del grupo (o un subgrupo de ellos) agreden al individuo inmigrante desplazándolo de los lugares de alimentación y tratando de evitar que se incorpore. Además, si la dispersión tiene lugar hacia un área desconocida, el individuo puede resultar más vulnerable frente al ataque de los depredadores y sufrir ciertos costes como los derivados del desconocimiento de la localización de los recursos alimenticios. Aunque existen estrategias para reducir estos costes, como la dispersión en compañía de familiares o el establecimiento de algunas relaciones con el grupo antes de incorporarse a él, la dispersión es claramente un proceso costoso para los individuos implicados, lo que nos lleva a plantearnos el porqué los individuos exhiben este comportamiento.
A la hora de intentar responder a esta cuestión hay que distinguir entre las causas proximales, que hacen referencia a los mecanismos implicados en la expresión del comportamiento, y las causas últimas, que están relacionadas con las consecuencias adaptativas del comportamiento y, por tanto, con la explicación de su selección y evolución. Entre las posibles causas proximales para la dispersión en primates se encuentran: la atracción por individuos de otros grupos, los factores fisiológicos, el estado nutricional del individuo, las tasas de interacción con los miembros del grupo (entre las que se incluirían las agresiones o las interacciones sexuales), la coerción o la respuesta a perturbaciones ambientales. En relación a las causas últimas de la dispersión en primates, se han sugerido cuatro grandes categorías. En primer lugar, los individuos podrían abandonar su grupo para evitar la competencia por los recursos tróficos con los miembros de este, especialmente con sus parientes. En segundo lugar, los individuos podrían abandonar su grupo en respuesta a la competencia intrasexual por las parejas reproductivas, especialmente la que ocurre entre parientes. En tercer lugar, las hembras podrían emplear la dispersión como una estrategia frente al infanticidio por parte de los machos. El infanticidio representa una estrategia adaptativa para ciertos machos no emparentados con las crías debido a que reduce el periodo de infertilidad posparto y acelera la tasa reproductiva de la hembra. Por último, los individuos que abandonan el grupo natal podrían obtener un beneficio al aparearse con individuos no emparentados y evitar los efectos negativos para la supervivencia de las crías derivados de los apareamientos endogámicos (i. e. depresión consanguínea).

En la imagen, dos langures Hanuman (Semnopithecus entellus) en la India. Esta especie de cercopitécidos asiáticos forma grupos de 15 a 54 individuos entre los que podemos encontrar un solo macho reproductor o varios. La dispersión es masculina y está asociada con la competencia por las parejas reproductivas. / Daniele Colombo
Características de los patrones de dispersión
¿Quién se dispersa?
En la mayoría de mamíferos existe un sesgo en el patrón de dispersión en relación con el sexo, siendo el macho el que generalmente abandona la unidad natal. Esto es debido a la asimetría en la inversión parental que se produce de manera general en los mamíferos (las hembras invierten más que los machos) y que afecta a las tasas reproductivas potenciales (número de crías que cada sexo podría producir por unidad de tiempo si su reproducción no se encontrase limitada por la disponibilidad de parejas reproductivas) y a los factores que limitan el éxito reproductivo en uno y otro sexo (Trivers, 1972). Por un lado, permanecer en el entorno natal permite a las hembras tener acceso a una fuente de alimento conocida, que representa el factor limitante para su reproducción y, a su vez, les facilita el establecimiento de una serie de relaciones cooperativas entre individuos emparentados (al dispersarse los machos, estas relaciones se establecerían entre hembras emparentadas formando las llamadas matrilíneas) y no emparentados que tiene un impacto positivo sobre su supervivencia y reproducción. Por otro lado, aunque los machos podrían beneficiarse igualmente de permanecer en su entorno natal, su éxito reproductivo se encuentra limitado por el número de parejas a las que tienen acceso, por lo que los costes de la dispersión podrían verse compensados al incorporarse a unidades en las que optimizasen dicho acceso.
«En la mayoría de mamíferos existe un sesgo en el patrón de dispersión en relación con el sexo y es el macho el que generalmente abandona la unidad natal»
En primates hay diversas especies que pueden incluirse dentro de este patrón general de dispersión (tabla 2). Un ejemplo de dispersión masculina asociada a la competencia por las parejas reproductoras lo encontramos en los langures Hanuman (figura 1) (Koenig y Borries, 2001). Esta especie, perteneciente a la familia de los cercopitécidos, puede formar grupos con un solo macho adulto y varias hembras (unimacho-multihembra) o grupos multimacho-multihembra. En el primer caso, los machos suelen abandonar el grupo alrededor de la madurez sexual, aunque pueden ser expulsados antes si el macho dominante (posiblemente su padre) es reemplazado por otro individuo ajeno al grupo. Por otra parte, en los grupos multimacho-multihembra, los individuos pueden permanecer durante más tiempo en sus unidades natales antes de dispersarse. En ambos casos, la competencia por las parejas reproductoras se postula como la principal causa de la dispersión. En el primer caso, un macho es capaz de expulsar al resto de competidores de la unidad y, en el otro, el macho dominante es el que monopoliza la mayor parte de la paternidad en el grupo. Los machos que se dispersan suelen unirse a grupos que solo contienen machos en espera de formar o acceder a un grupo reproductor.
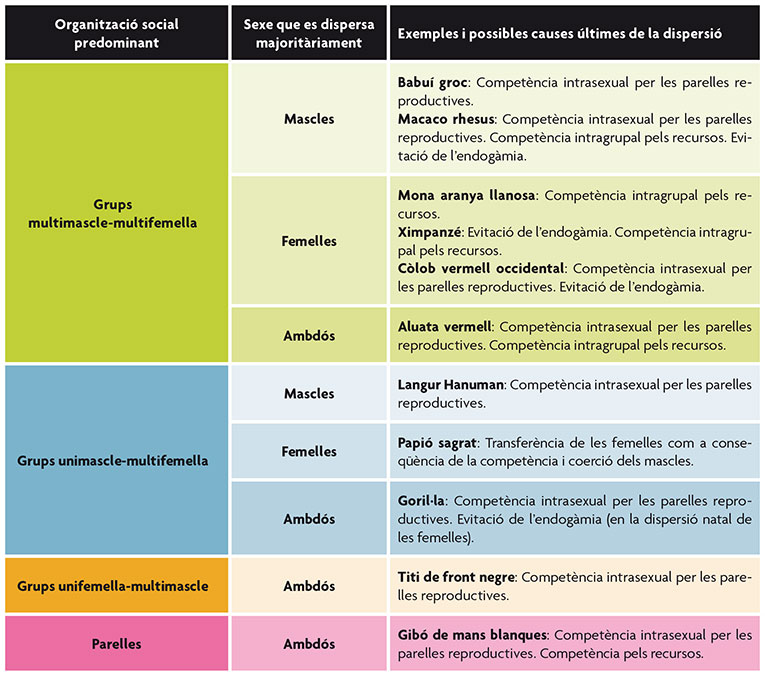
Tabla 2. Principales patrones de dispersión descritos en primates en función del tipo predominante de organización social de la especie y del sesgo sexual en la dispersión. A su vez, se muestran varios ejemplos y las causas últimas más probables de la dispersión en cada caso.
Sin embargo, existen diversas especies que no se ajustan a este patrón general de dispersión y en las que las hembras habitualmente abandonan sus unidades natales mientras que los machos permanecen en ellas (tabla 2). Se considera que la competencia por los recursos tróficos constituiría la causa más frecuente que determinaría este patrón, aunque la evitación de la endogamia o la elección de pareja también constituyen causas probables en ciertos casos. Los muriquis son una especie de monos del Nuevo Mundo, pertenecientes a la familia de los atélidos, que forman grupos multimacho-multihembra con un sistema de fusión-fisión en el que las hembras abandonan habitualmente el grupo natal alrededor de la madurez sexual. Aunque no son expulsadas agresivamente del grupo, las hembras adultas las desplazan con frecuencia de los sitios de alimentación, por lo que se piensa que dicha dispersión se debe en gran medida a la competencia por los recursos tróficos (Printes y Strier, 1999). Los chimpancés, pertenecientes a la familia de los homínidos, también presentan sistemas sociales de fusión-fisión formados por grupos multimacho-multihembra (Stumpf, 2007). Los machos son filopátricos, ya que permanecen en sus comunidades natales, en las que forman coaliciones y cooperan en la defensa del territorio, siendo las hembras las que se dispersan a otros grupos alrededor de la madurez sexual. Sin embargo, a diferencia de lo que ocurre en los muriquis, esta dispersión parece estar más relacionada con la atracción de las hembras hacia los machos de fuera del grupo que con la competición por el alimento dentro del grupo. En este caso, las causas proximales de la dispersión parecen indicar que ésta estaría mayoritariamente relacionada con la evitación de la endogamia, aunque la competencia por los recursos tróficos podría estar jugando cierto papel en el proceso. Por último, la especie humana también representa una excepción al patrón general de dispersión en los primates, ya que, a pesar de que ambos sexos pueden abandonar sus unidades natales, la dispersión se encuentra sesgada hacia las hembras en la mayoría de las sociedades estudiadas (Rodseth et al., 1991).
En otras especies no existe un sesgo sexual en el patrón de dispersión, ya que ambos sexos abandonan el grupo natal (tabla 2). Los gorilas, pertenecientes a la familia de los homínidos, constituyen un ejemplo de este patrón de dispersión (figura 2). Sus grupos sociales están formados por un solo macho reproductor o, en ocasiones, por dos, aunque el dominante acapara la mayoría de los apareamientos y, por tanto, de la paternidad en el grupo (Harcourt y Stewart, 2007). Consecuentemente, la principal causa de dispersión en los machos sería la competencia reproductiva. En el caso de las hembras, esta dispersión está relacionada con la evitación de los apareamientos endogámicos y con la elección de pareja.
«El infanticidio representa una estrategia adaptativa para ciertos machos no emparentados con las crías debido a que reduce el periodo de infertilidad posparto»
Para finalizar, es preciso señalar que el tipo de sesgo sexual en la dispersión constituye un factor importante, ya que determinará la estructura de parentesco dentro de los grupos e influirá sobre las características de las relaciones sociales (i. e. estructura social). En general, el sexo que se dispersa pierde la mayor parte de las relaciones sociales con sus parientes.
A su vez, los vínculos que establece en el nuevo grupo con los individuos del mismo sexo suelen ser débiles o inexistentes. En contraposición, el sexo filopátrico, el que permanece en su entorno natal, establece fuertes vínculos cooperativos y alianzas con sus parientes formando el núcleo de la estructura social del grupo (matrilíneas o patrilíneas, dependiendo del sexo). De esta manera, filopatría, consanguinidad y cooperación tienden a coincidir. Sin embargo, en la especie humana, las hembras tienden a mantener los vínculos con sus parientes, a pesar de separarse de ellos, excepto en sociedades con un componente patriarcal extremo.
A su vez, también pueden formar vínculos cooperativos en sus nuevos grupos con hembras con las que no están emparentadas, circunstancia que se da en menor medida en otras especies de primates con dispersión femenina con excepción, quizá, de los bonobos. Por otro lado, en aquellas sociedades humanas en las que las hembras son filopátricas, los machos tampoco pierden los vínculos con sus parientes y, de hecho, estas sociedades suelen estar caracterizadas por un alto grado de endogamia comunitaria, ya que los machos o no abandonan la comunidad natal o se desplazan a grupos vecinos, muchas veces dependientes de la comunidad natal (Rodseth et al., 1991).
¿Cuándo dispersarse? El momento de la dispersión y su relación con el infanticidio
Podemos hablar de dos tipos de dispersión social en función del momento en el que tiene lugar. La dispersión primaria (o natal) es la que ocurre alrededor de la madurez sexual y supone una separación del individuo de su grupo natal, mientras que la dispersión secundaria constituye un cambio de unidad social durante la edad adulta.
«la especie humana representa una excepción al patrón general de dispersión en los primates, ya que, la dispersión se encuentra sesgada hacia las hembras en la mayoría de las sociedades estudiadas»
El momento en el que los individuos se dispersan de su unidad natal puede ser crítico para su supervivencia, sin embargo, este es un parámetro que varía sustancialmente entre especies y para diferentes individuos dentro de una misma especie, como hemos visto en el caso de los langures Hanuman. Se piensa que el momento de la dispersión está determinado por el balance entre los costes potenciales de permanecer en el grupo (depresión consanguínea, agresión por parte de los miembros del grupo, acceso limitado a los recursos tróficos y sociales) y los costes potenciales de abandonarlo, que han sido descritos previamente.
En algunas especies en las que las hembras se dispersan secundariamente, la decisión de cuándo hacerlo se encuentra influida por el riesgo de infanticidio (Sterck y Korstjens, 2000). En primer lugar, por norma general, las hembras cambian de grupo social cuando no tienen crías dependientes a su cargo para evitar agresiones infanticidas por parte de los machos en los nuevos grupos. Y en segundo lugar, en aquellas especies, como los gorilas, los langures de Thomas o los colobos rojos, en las que la paternidad se concentra en un solo macho o está fuertemente sesgada hacia él, la percepción del riesgo de que este macho sea reemplazado por otro externo al grupo, lo que ocasionaría una situación de riesgo de infanticidio para las crías, es el factor que desencadena la dispersión de las hembras no lactantes, en ocasiones, al grupo del potencial macho infanticida. Esta dispersión se interpreta desde el punto de vista de la elección de pareja, ya que las hembras parece que cambian de unidad social en busca de un macho que les proporcione una mejor protección para sus crías frente a las agresiones.
El modo de dispersión. Conflicto sexual y dispersión en los babuinos hamadríades
«La dispersión se puede producir por iniciativa propia o como consecuencia del rechazo y agresión por parte de los individuos del grupo»
Como ya hemos adelantado, la dispersión se puede producir por iniciativa propia o como consecuencia del rechazo y agresión por parte de los individuos del grupo. Sin embargo, en los babuinos hamadríades, pertenecientes a la familia de los cercopitécidos, las hembras no se dispersan por ninguno de estos dos procesos sino que son «secuestradas». El secuestro es un tipo de coerción sexual que implica la separación forzosa de una hembra de su unidad social. En los babuinos hamadríades este es un proceso complejo en el que los machos compiten por la hembra y esta, a su vez, recibe una agresión intensa por parte del macho secuestrador cuya función es la de condicionar el comportamiento de la hembra a cambiar de unidad y que tiene asociado un coste para el éxito reproductivo inmediato de la hembra (Polo y Colmenares, 2012; Polo et al., 2014) (figura 3). De esta manera, la dispersión de las hembras entre las unidades unimacho, que constituye la unidad social y reproductiva básica en esta especie, está más relacionada con una estrategia coercitiva del macho que con una estrategia reproductiva de la hembra, ya que el cambio de grupo está forzado por el macho y es costoso para la hembra.

En los babuinos hamadríades (Papio hamadryas hamadryas) los machos agreden con intensidad a las hembras que están siendo transferidas a sus unidades. El mordisco en la nuca es una de las agresiones características que se produce durante estos episodios denominados pastoreos. / Pablo Polo
Consideraciones finales
A medida que los patrones de dispersión se caracterizan mejor y para un número mayor de especies, lo que se observa es que existen numerosas excepciones a los patrones generales que se han descrito. En primer lugar, aunque la causa última de la dispersión de los machos se relaciona en muchas especies con la competencia intragrupal por el acceso a las parejas reproductivas, también hay casos en los que dicha causa parece estar más relacionada con la competencia por el alimento dentro del grupo o la evitación de la endogamia. A su vez, la dispersión en las hembras parece estar más relacionada con la competencia por el alimento dentro del grupo o la evitación de la endogamia, no obstante, en ciertas especies se piensa que las hembras también podrían abandonar su grupo en busca de mejores oportunidades reproductivas. De hecho, según el consenso actual, se considera que la dispersión en primates aparece como resultado de la combinación de estas causas o de algunas de ellas, tanto a nivel interespecífico como intraespecífico. Y, en segundo lugar, no todos los individuos de una población se comportan según el patrón general de dispersión de esta. Por ejemplo, en algunos casos, las hembras de chimpancés no se dispersan y permanecen durante toda su vida en los grupos natales. A su vez, se han descrito casos en especies donde las hembras constituyen el sexo filopátrico y en las que algunas de ellas abandonan su grupo natal (por ejemplo, en los monos capuchinos). Estos casos se pueden explicar desde una aproximación al fenómeno de la dispersión basada en el individuo, debido a que los costes y beneficios de la dispersión no son solo dependientes de la especie y del sexo, sino que también lo son de la situación de cada individuo en el grupo (Jack e Isbell, 2009). De manera que, para ciertas hembras, en especies con filopatría femenina, el balance entre costes y beneficios puede favorecer el abandono del grupo debido, por ejemplo, a una baja posición en la jerarquía de dominancia del grupo que limitaría su acceso a los recursos. Estas dos características que hemos descrito provocan la aparición de la gran variabilidad tanto interespecífica como intraespecífica e interindividual que encontramos en los patrones de dispersión en los primates.
Field, M. i D. Guatelli-Steinberg, 2003. «Dispersal and the Inbreeding Avoidance Hypothesis». Primate Report, 67: 7-60.
Harcourt, A. H. i K. J. Stewart, 2007. Gorilla Society: Compromise and Cooperation between Sexes. University of Chicago Press. Chicago.
Isbell, L. A. i D. Van Vuren, 1996. «Differential Costs of Locational and Social Dispersal and Their Consequences for Female Group-living Primates». Behaviour, 133: 1-36. DOI: <10.1163/156853996X00017>.
Jack, K. M. i L. A. Isbell, 2009. «Dispersal in Primates: Advancing An Individualized Approach». Behaviour, 146: 429-436. DOI: <10.1163/156853909X410612>.
Koenig, A. i C. Borries, 2001. «Socioecology of Hanuman Langurs: the Story of Their Success». Evolutionary Anthropology: Issues, News, and Reviews, 10: 122-137. DOI: <10.1002/evan.1026>.
Polo, P. i F. Colmenares, 2012. «Behavioural Processes in Social Context: Female Abductions, Male Herding and Female Grooming in Hamadryas Baboons». Behavioural Processes, 90: 238-245. DOI: <10.1016/j.beproc.2012.02.004>.
Polo, P.; Hernández-Lloreda, M. V. i F. Colmenares, 2014. «Male Takeovers Are Reproductively Costly to Females in Hamadryas Baboons: A Test of The Sexual Coercion Hypothesis». PLOS ONE, 9(3): e90996. DOI: <10.1371/journal.pone.0090996>.
Printes, R. C. i K. B. Strier, 1999. «Behavioral Correlates of Dispersal in Female Muriquis (Brachyteles arachnoides)». International Journal of Primatology, 20: 941-960. DOI: <10.1023/a:1020882719850>.
Rodseth, L. et al., 1991. «The Human Community as a Primate Society [and Comments]». Current Anthropology, 32: 221-254. DOI: <10.2307/2743773>.
Sterck, E. H. M. i A. H. Korstjens, 2000. «Female Dispersal and Infantice Avoidance in Primates». En Van Schaik, C. P. i C. H. Janson (eds.). Infanticide by Males and its Implications. Cambridge University Press. Cambridge.
Stumpf, R., 2007. «Chimpanzees and Bonobos: Diversity within and between Species». En Campbell, C. J.; Fuentes, A.; MacKinnon, K. C.; Panger, M. y S. K. Bearder (eds.). Primates in Perspective. Oxford University Press. Oxford.
Trivers, R., 1972. «Parental Investment and Sexual Selection». En Campbell, B. (ed.). Sexual Selection and The Descent of Man 1871-1971. Aldine-Atherton. Chicago.




