Biodiversidad marina en el tiempo y el espacio
Lo que nos cuentan los fósiles minúsculos

La biodiversidad ha ido cambiando tanto en el espacio como en el tiempo. Por ejemplo, encontramos más especies en los trópicos y menos en la región ártica y la antártica, siguiendo un gradiente latitudinal de diversidad, uno de los patrones que podemos observar de manera más consistente en este mundo complejo. Lo que sabemos sobre los gradientes temporales de biodiversidad es mucho más limitado. Esto ocurre porque haría falta un programa de seguimiento continuo bien diseñado, y estos difícilmente persisten más allá de unas décadas. Pero, por suerte, tenemos restos de antiguos organismos: los fósiles. Los fósiles son básicamente el único registro directo de la biodiversidad pasada.
Palabras clave: microfósiles, ecología, evolución, paleontología, biogeografía.
«Pequeños organismos conocidos como ‘microfósiles’ nos permiten inferir patrones de biodiversidad pasados de forma detallada»
La belleza y utilidad de los fósiles minúsculos
Algunos organismos tienen registros fósiles extraordinarios. En especial, los pequeños organismos conocidos como microfósiles (fósiles de tamaño microscópico) nos permiten inferir de forma detallada patrones de biodiversidad pasados, puesto que podemos encontrar decenas de miles de ellos en un solo puñado de sedimentos y rocas (Yasuhara, Tittensor, Hillebrand y Worm, 2017). Por ejemplo, conocemos el gradiente latitudinal de diversidad de varios organismos marinos (Figura 1) (Hillebrand, 2004), pero no es fácil saber cómo era el gradiente en el pasado porque, por ejemplo, los peces (o la mayoría del resto de organismos) rara vez se conservan como fósiles. Utilizando un grupo de microfósiles, los procedentes de foraminíferos planctónicos (Figura 2), podemos observar la biodiversidad del pasado y su gradiente latitudinal en el piélago oceánico.
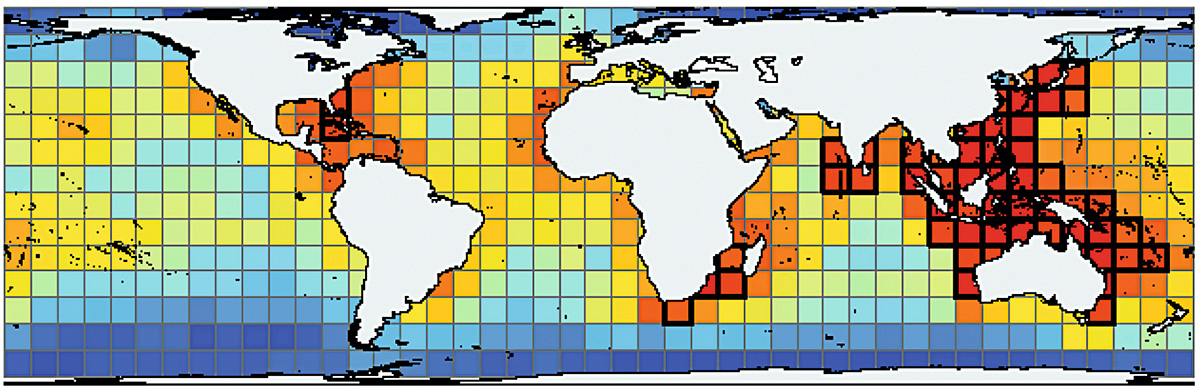
Figura 1. Patrón mundial de biodiversidad marina. Los colores más cálidos indican mayor biodiversidad (más especies). / © Tittensor et al. (2010)
Puede que los foraminíferos planctónicos (protozoos con concha que suelen medir entre 0,1 y 1 mm) no sean famosos, pero son unos microfósiles muy importantes por varias razones. En primer lugar, se conservan millones de ellos en sedimentos y rocas y suponen un registro geológico y paleontológico casi perfecto. En segundo lugar, casi todas las especies cenozoicas se han descrito formalmente, su identificación estandarizada está bien establecida por su importancia en la bioestratigrafía y la paleoceanografía, y el número de especies es manejable (no son demasiado numerosas: se conocen unas 40 especies planctónicas en total en los océanos actuales); en otras palabras, su taxonomía es robusta (Yasuhara, Hunt, Dowsett, Robinson y Stoll, 2012; Yasuhara, Tittensor et al., 2017).
«Cada vez es más importante integrar los datos biológicos y los fósiles para comprender mejor nuestro planeta»
Al comparar los patrones latitudinales de diversidad entre el presente y la última glaciación (hace 20.000 años), observamos la persistencia del gradiente que se extiende desde los trópicos hasta los polos (Yasuhara et al., 2012) (Figura 3). Pero analizando los detalles está claro que el gradiente de diversidad de la glaciación es mucho más pronunciado que el presente. Aunque las diversidades tropicales son similares entre la última glaciación y la actualidad, las diversidades de ambas empiezan a divergir a partir de 20–30º N y hacia latitudes mayores (Figura 3a). ¿Por qué? Si comparamos las diversidades no solo con la latitud sino también con la temperatura del océano, encontramos una posible respuesta. Conocemos bien la temperatura superficial del océano durante la última glaciación gracias a diferentes métodos micropaleontológicos y geoquímicos (Yasuhara et al., 2012). La relación entre temperatura y diversidad es sorprendentemente consistente entre la actualidad y la última glaciación (Figura 3b). Por lo tanto, el gradiente latitudinal de diversidad era más pronunciado durante la última glaciación porque el frío severo en las latitudes altas acentuaba el gradiente latitudinal de temperatura (Yasuhara et al., 2012).
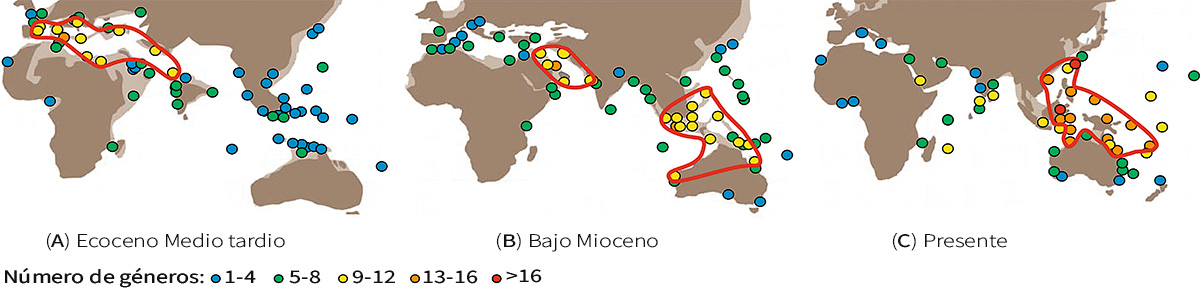
Figura 5. Distribución de la diversidad global de los géneros de grandes foraminíferos bentónicos (A) en el Eoceno Medio tardío (hace 42–39 millones de años), (B) en el Bajo Mioceno (hace 23–16 millones de años), y (C) en el presente. Se puede observar que los puntos calientes de diversidad (señalados con líneas rojas) han ido variando. En el Eoceno, la diversidad más alta se encontraba en el océano de Tetis (el antiguo Mediterráneo), mientras que en el Bajo Mioceno, se encontraba en la zona de la península de Arabia. El punto caliente del triángulo de coral del archipiélago indoaustraliano también empezó a formarse en esta época, hace unos veinte millones de años. Actualmente, esta zona sigue siendo un punto caliente de biodiversidad. / © Renema et al. (2008)
Puntos calientes de biodiversidad cambiantes
Incluso en los océanos tropicales, a los que se reconoce como regiones biodiversas, la biodiversidad no permanece estable en términos espaciales ni temporales. Algunas áreas tienen muchas más especies que otras. La diversidad de especies no varía solo con la latitud, como he explicado en la sección anterior, sino también con la longitud. El triángulo de coral es un ejemplo de ello. Está caracterizado por la mayor biodiversidad marina del mundo (Figura 1 y 4) (Bellwood, Renema y Rosen, 2012), pero incluso su diversidad ha cambiado con el tiempo.
«Incluso en regiones biodiversas como los océanos tropicales, la biodiversidad no permanece estable en términos espaciales ni temporales»
Los grandes foraminíferos (Figura 2), un tipo de foraminíferos bentónicos con grandes conchas y algas simbióticas, muestran evidencia directa de que la biodiversidad del triángulo de coral ha cambiado. Quizás no nos sorprenda que la diversidad en este área marítima se haya mantenido persistentemente alta en los últimos veinte millones de años, pero, antes de eso, la biodiversidad de la región era más modesta. En el Eoceno, por ejemplo, la diversidad más alta se encontraba en el océano de Tetis, el antiguo Mediterráneo (Figura 5) (Renema et al., 2008; Yasuhara, Tittensor et al., 2017).
«Los microfósiles nos ayudan a entender mejor los ecosistemas pasados, presentes y futuros»
El punto caliente de biodiversidad del triángulo de coral se estableció hace cerca de veinte millones de años, como también indican los fósiles y los datos moleculares de otros organismos como los ostrácodos, los gasterópodos y los peces (Renema et al., 2008; Yasuhara, Iwatani, et al., 2017). Es probable que este origen del punto caliente de biodiversidad del triángulo de coral esté relacionado con la colisión de Australia contra los arcos del Pacífico y el sudeste asiático hace entre veintitrés y veinticinco millones de años, que formó grandes hábitats marinos superficiales, diversos, complicados y dinámicos, lo que ayudó a la biodiversidad de la región (Renema et al., 2008; Yasuhara, Iwatani et al., 2017; Yasuhara, Tittensor et al., 2017).

Figura 3. Gradiente latitudinal de diversidad de especies para los foraminíferos planctónicos en la actualidad y durante la última glaciación (hace 20.000 años). Las líneas son curvas suavizadas del diagrama de dispersión y nos permiten visualizar una relación constante entre la temperatura del océano y la biodiversidad de foraminíferos planctónicos, aunque el gradiente de diversidad de la glaciación es más pronunciado que el actual. Esto podría deberse al frío severo en las latitudes altas durante la última glaciación en comparación con la temperatura actual. Modificada a partir de Yasuhara et al. (2012).
Clima y biodiversidad de aguas profundas
Las aguas profundas son otro ecosistema caracterizado por albergar numerosas especies (Ingels et al., 2016; Rex y Etter, 2010). El impacto climático en la biodiversidad de aguas profundas es un tema que solo se puede tratar de forma adecuada si utilizamos los registros paleontológicos, especialmente los de microfósiles, porque, en primer lugar, nuestro seguimiento de los organismos marinos modernos no suele ser lo suficientemente prolongado para poder evaluar los efectos de los cambios climáticos que ocurren en períodos de tiempo más largos que la vida humana (Yasuhara, Doi, Wei, Danovaro y Myhre, 2016); y, segundo, porque la mayoría de los organismos de aguas profundas, especialmente los grandes, no tienen estructuras mineralizadas (los bivalvos y los gasterópodos tienen conchas pero su abundancia fósil es demasiado baja en los sedimentos de aguas profundas) y, por tanto, apenas tienen registro fósil.
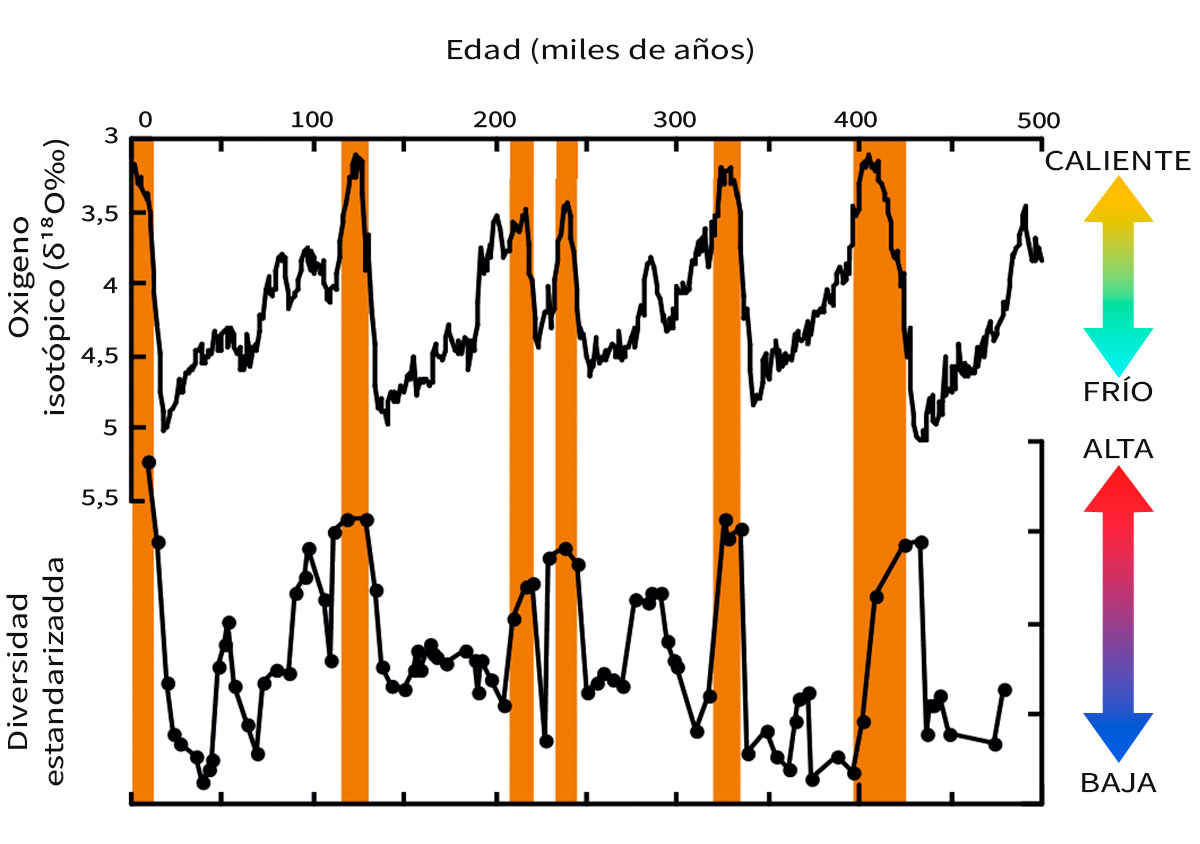
Figura 6. Comparación entre los cambios en el clima global y en la diversidad de especies de ostrácodos de aguas profundas durante los últimos 500.000 años. La curva isotópica del oxígeno representa los cambios climáticos globales, y los valores isótopicos más bajos indican intervalos más cálidos. Los picos principales de clima cálido están marcados con barras naranjas. Se puede observar que la biodiversidad de estas especies de ostrácodos es mayor en épocas más cálidas y menor durante las glaciaciones. Modificada a partir de Yasuhara et al. (2009)
Los sedimentos depositados en el fondo del océano son un archivo ideal para estudiar este tema porque contienen numerosos microfósiles, además de registros climáticos excelentes que se pueden reconstruir mediante varios métodos geoquímicos. Los microfósiles de bellos crustáceos pequeños (por ejemplo, ostrácodos como los que muestra la Figura 2, que en general miden aproximadamente 1 mm o incluso menos y son muy sensibles a los cambios ambientales, con más de 20.000 especies vivas estimadas) muestran una clara correlación con los cambios climáticos glaciales e interglaciares (Cronin y Raymo, 1997; Yasuhara y Cronin, 2008; Yasuhara, Hunt, Cronin y Okahashi, 2009). Su biodiversidad es mayor en épocas más cálidas y menor en las glaciaciones (Figura 6) (Yasuhara et al., 2009). Conocemos bien este control térmico de la biodiversidad de aguas profundas a diferentes escalas, desde el presente, hasta décadas o cientos de miles de años (Yasuhara y Danovaro, 2016).
Salvando las distancias
Tradicionalmente, las diferentes comunidades de investigación han estudiado la biología y la paleontología por separado. Sin embargo, cada vez es más importante integrar los datos biológicos y los fósiles para comprender mejor nuestro planeta y los organismos que viven en él (Yasuhara, Tittensor et al., 2017). Nuestro planeta está cambiando rápidamente debido a diferentes actividades humanas, y conocer las respuestas pasadas a cambios similares es muy importante para que podamos prepararnos mejor para el futuro. Gracias a su extraordinario registro fósil, los microfósiles, aunque pequeños en tamaño, tienen un papel crucial en la respuesta a este problema.
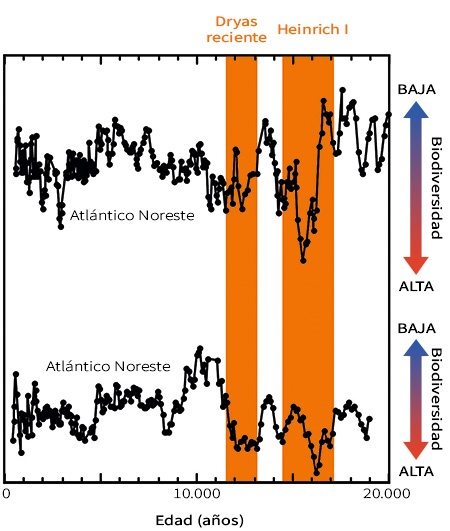
Figura 7. Cambios en la biodiversidad de aguas profundas en el noreste y el noroeste del océano Atlántico durante los últimos 20.000 años. Los registros fósiles de estas localizaciones tan distantes (50.000 km) muestran aumentos de biodiversidad rápidos y sincrónicos durante los sucesos de cambio climático rápido del Dryas reciente y el Heinrich I (marcados en naranja). Mayor biodiversidad significa más especies en una muestra. Modificada a partir de Yasuhara et al. (2014)
Como observación final, resulta notable lo poco que sabemos sobre cómo responde la biodiversidad marina a los cambios climáticos rápidos en un entorno a lo largo de décadas o siglos. Estas escalas temporales son relevantes para los cambios antropogénicos presentes y futuros, como el calentamiento global, la desoxigenación, la acidificación, la contaminación, el aumento del nivel del mar y la sobrepesca (Breitburg et al., 2018; Jackson et al., 2001; Mora et al., 2013). Estas escalas de tiempo han sido un «punto ciego» para los estudios biológicos y paleoecológicos, que son demasiado largos para la observación humana y demasiado cortos para resolverse con los registros fósiles ordinarios. Gracias a registros de microfósiles excepcionalmente detallados podemos estudiar estas escalas de tiempo. De hecho, diferentes estudios recientes que han utilizado registros de microfósiles han mostrado, por ejemplo, que la biodiversidad de aguas profundas ha sido sensible a cambios climáticos abruptos a estas escalas temporales (Yasuhara et al., 2016; Yasuhara, Cronin, DeMenocal, Okahashi y Linsley, 2008; Yasuhara, Okahashi, Cronin, Rasmussen y Hunt, 2014). Los registros de microfósiles mostraban evidentes marcas en la biodiversidad de aguas profundas debido a sucesos conocidos de cambio climático abrupto, como el Heinrich I (hace 17.000–14.600 años) y el Dryas Reciente (hace 12.900–11.700 años) (Figura 7) (Yasuhara et al., 2014). Los modelos estadísticos indicaban que estos cambios en la diversidad fueron causados probablemente por el calentamiento de las aguas profundas a nivel regional durante estos sucesos de cambio climático abrupto (Yasuhara et al., 2014). Estos cambios podrían haber afectado no solo a la biodiversidad, sino también al funcionamiento de los ecosistemas de aguas profundas (Yasuhara et al., 2016). Los microfósiles nos ayudan a entender mejor los ecosistemas pasados, presentes y futuros, salvando la distancia que separa la escala temporal biológica y la paleontológica (Mesquita-Joanes, Smith y Viehberg, 2012; Yasuhara, Tittensor et al., 2017).
Referencias
Bellwood, D. R., Renema, W., & Rosen, B. R. (2012). Biodiversity hotspots, evolution and coral reef biogeography: A review. En D. J. Gower, K. G. Johnson, J. E. Richardson, B. R. Rosen, L. Rüber, & S. T. Williams (Eds.), Biotic evolution and environmental change in Southeast Asia (pp. 216–245). Cambridge: Cambridge University Press.
Breitburg, D., Levin, L. A., Oschlies, A., Grégoire, M., Chavez, F. P., Conley, D. J., … Zhang, J. (2018). Declining oxygen in the global ocean and coastal waters. Science, 359(6371), eaam7240. doi: 10.1126/science.aam7240
Cronin, T. M., & Raymo, M. E. (1997). Orbital forcing of deep-sea benthic species diversity. Nature, 385, 624–627. doi: 10.1038/385624a0
Hillebrand, H. (2004). Strength, slope and variability of marine latitudinal gradients. Marine Ecology Progress Series, 273, 251–267. doi: 10.3354/meps273251
Ingels, J., Clark, M. R., Vecchione, M., Perez, J. A. A., Levin, L. A., Priede, I. G., … Van Gaever, S. (2016). Chapter 36F. Open Ocean Deep Sea. En L. Inniss & Simcock (Eds.), First global integrated marine assessment. World ocean assessment I. Nova York, NY: Organización de las Naciones Unidas.
Jackson, J. B. C., Kirby, M. X., Berger, W. H., Bjorndal, K. A., Botsford, L. W., Bourque, B. J., … Warner, R. R. (2001). Historical overfishing and the recent collapse of coastal ecosystems. Science, 293(5530), 629-638. doi: 10.1126/science.1059199
Mesquita-Joanes, F., Smith, A. J., & Viehberg, F. A. (2012). The ecology of Ostracoda across levels of biological organisation from individual to ecosystem: A review of recent developments and future potential. En D. J. Horne, J. Holmes, J. Rodriguez-Lazaro, & F. A. Viehberg (Eds.), Ostracoda as proxies for Quaternary climate change (pp. 15–35). Amsterdam: Elsevier.
Mora, C., Wei, C. L., Rollo, A., Amaro, T., Baco, A. R., Billett, D., ... Yasuhara, M. (2013). Biotic and human vulnerability to projected changes in ocean biogeochemistry over the 21st century. PLoS Biology, 11(10), e1001682. doi: 10.1371/journal.pbio.1001682
Renema, W., Bellwood, D. R., Braga, J. C., Bromfield, K., Hall, R., Johnson, K. G., … Pandolfi, J. M. (2008). Hopping hotspots: Global shifts in marine biodiversity. Science, 321(5889), 654–657. doi: 10.1126/science.1155674
Rex, M. A., & Etter, R. J. (2010). Deep-sea biodiversity: Pattern and scale. Cambridge: Harvard University Press.
Tittensor, D. P., Mora, C., Jetz, W., Lotze, H. K., Ricard, D., Berghe, E. V., & Worm, B. (2010). Global patterns and predictors of marine biodiversity across taxa. Nature, 466, 1098-1101. doi: 10.1038/nature09329
Yasuhara, M., & Cronin, T. M. (2008). Climatic influences on deep-sea ostracode (Crustacea) diversity for the last three million years. Ecology, 89(11), S53–S65. doi: 10.1890/07-1021.1
Yasuhara, M., Cronin, T. M., DeMenocal, P. B., Okahashi, H., & Linsley, B. K. (2008). Abrupt climate change and collapse of deep-sea ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 105(5), 1556–1560. doi: 10.1073/pnas.0705486105
Yasuhara, M., & Danovaro, R. (2016). Temperature impacts on deep-sea biodiversity. Biological Reviews, 91(2), 275–287. doi: 10.1111/brv.12169
Yasuhara, M., Doi, H., Wei, C. L., Danovaro, R., & Myhre, S. E. (2016). Biodiversity-ecosystem functioning relationships in long-term time series and palaeoecological records: Deep sea as a test bed. Philosophical Transactions of the Royal Society B, 371(1694). doi: 10.1098/rstb.2015.0282
Yasuhara, M., Hunt, G., Cronin, T. M., & Okahashi, H. (2009). Temporal latitudinal-gradient dynamics and tropical instability of deep-sea species diversity. Proceedings of the National Academy of Sciences of the United States of America, 106(51), 21717–21720. doi: 10.1073/pnas.0910935106
Yasuhara, M., Hunt, G., Dowsett, H. J., Robinson, M. M., & Stoll, D. K. (2012). Latitudinal species diversity gradient of marine zooplankton for the last three million years. Ecology Letters, 15(10), 1174–1179. doi: 10.1111/j.1461-0248.2012.01828.x
Yasuhara, M., Iwatani, H., Hunt, G., Okahashi, H., Kase, T., Hayashi, H., … Renema, W. (2017). Cenozoic dynamics of shallow-marine biodiversity in the Western Pacific. Journal of Biogeography, 44(3), 567–578. doi: 10.1111/jbi.12880
Yasuhara, M., Okahashi, H., Cronin, T. M., Rasmussen, T. L., & Hunt, G. (2014). Response of deep-sea biodiversity to abrupt deglacial and Holocene climate changes in the North Atlantic Ocean. Global Ecology and Biogeography, 23(9), 957–967. doi: 10.1111/geb.12178
Yasuhara, M., Tittensor, D. P., Hillebrand, H., & Worm, B. (2017). Combining marine macroecology and palaeoecology in understanding biodiversity: Microfossils as a model. Biological Reviews, 92(1), 199–215. doi: 10.1111/brv.12223