

Saurópodos virtuales
Una revolución en el estudio de los dinosaurios de cuello largo
https://doi.org/10.7203/metode.14.24689

El estudio de los fósiles mediante técnicas de visualización en tres dimensiones, mejor conocido como paleontología virtual, ha supuesto una revolución en la investigación de los organismos del pasado en los últimos veinte años. El estudio de los populares dinosaurios saurópodos es ejemplo de ello: los complejos sistemas de soportes o las costosas réplicas usados antaño para estudiar sus esqueletos son ahora realizados fácilmente gracias a los avances en computación. Cuestiones como la posición de sus largos cuellos, sus capacidades de movimiento e implicaciones funcionales o incluso la cópula son más fáciles de abordar que nunca. Así, evidencias obtenidas mediante la metodología tradicional junto con las aportaciones de las nuevas tecnologías abren nuevas fronteras en el estudio de estos colosos.
Palabras clave: paleontología virtual, rango de movimiento del cuello, cópula, jirafas, Jurásico.
Introducción: un revulsivo para la paleontología
Los saurópodos eran dinosaurios con un característico plan corporal: cuadrúpedos de largas colas y cuellos que adquirieron enormes tamaños. Los más grandes llegaron a alcanzar la altura de edificios de cuatro pisos y masas de hasta 70 toneladas (tanto como 250 elefantes), lo que los convierte en los animales más grandes que jamás hayan caminado sobre la Tierra (Hallett y Wedel, 2016; Sander et al., 2010). Cómo se desenvolvían estos animales tan impresionantes en sus ambientes pretéritos ha sido interpretado de múltiples maneras desde que los primeros esqueletos razonablemente completos fueron descubiertos a finales del siglo XIX, en un debate que se ha prolongado hasta el presente (Taylor, 2010). Considerados durante décadas como animales obligatoriamente acuáticos debido a su gran tamaño, pasaron a ser interpretados a finales del siglo XX como jirafas reptilianas: animales fitófagos (comedores de plantas) con cuellos imponentes coronados por pequeñas cabezas que se alimentaban de las copas de los árboles (Bakker, 1971). Sin embargo, el estudio de los fósiles a partir de visualización 3D, mejor conocido como paleontología virtual, ha supuesto un revulsivo no solo en el estudio de los fósiles en general (Sutton et al., 2014), sino también en la interpretación de los dinosaurios saurópodos desde principios de la década de los 2000 (Stevens, 2013; Stevens y Parrish, 1999; Taylor et al., 2009; Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020).
El debate sobre la posición del cuello
Los rápidos avances en tecnología computacional han permitido obtener información sin precedentes de los fósiles de dinosaurios. En el caso de los dinosaurios saurópodos, esta tecnología nos ha permitido desde estudiar el interior de las cavidades de sus huesos (como las de su sistema nervioso central o el oído) –sin necesidad de alterarlos físicamente gracias a la tomografía computerizada– hasta algo tan simple pero revolucionario como poder trabajar en un entorno virtual simultáneamente con decenas de huesos que en el mundo físico pesan cientos de kilos cada uno y resultan muy complicados de manejar.
En sus inicios, esta tecnología hizo posible un estudio que comparó los cuellos de los populares saurópodos Apatosaurus y Diplodocus para medir su rango de movimiento (Stevens y Parrish, 1999). El estudio digitalizó las facetas articulares de las vértebras cervicales de estos dinosaurios a partir de las ilustraciones de los huesos publicadas en las descripciones monográficas de estos taxones y calculó cuánto movimiento eran capaces de ejercer. Los resultados mostraban que ni Apatosaurus ni Diplodocus tenían cuellos tan flexibles como el de una jirafa actual, ni serían capaces de elevar sus cuellos muy por encima de sus hombros (Stevens y Parrish, 1999). Estudios posteriores parecían corroborar esta hipótesis, mostrando además que la columna vertebral de muchas especies de dinosaurios saurópodos se articulaba prácticamente recta, sin una elevación del cuello en su base como ocurre en muchos dinosaurios terópodos (Stevens y Parrish, 2005).
La conclusión estaba clara: esa imagen de los dinosaurios saurópodos como jirafas reptilianas que se alimentaban de las copas de los árboles debía ser revisada. Además, nuevos hallazgos de fósiles como el pequeño Nigersaurus de hace 110 millones de años mostraban claras adaptaciones a una alimentación de vegetación a nivel del suelo, como un cráneo cuyo hocico estaba orientado hacia abajo, extremidades anteriores cortas o una batería dental con decenas de dientes de reemplazo (Sereno et al., 2007). Estos hallazgos pronto influyeron en cómo se interpretaba la gran mayoría de saurópodos: animales con cuellos horizontales muy poco flexibles, casi incapaces de mirar lo que tenían a su espalda ni de levantar el cuello, lo que dio como resultado representaciones como la de Diplodocus en el célebre documental de 1999 Walking with dinosaurs de la BBC.
Sin embargo, esta hipótesis tan extendida de saurópodos de cuellos poco flexibles y más o menos horizontales no fue unánimemente aceptada por la comunidad científica. Se constató que los vertebrados terrestres actuales, desde los conejos a los camellos o los cocodrilos, habitualmente sostienen su cuello en posturas más elevadas de lo que sugerían los datos de la paleontología virtual (Taylor et al., 2009). ¿Quiere esto decir que los modelos virtuales no permiten inferir las capacidades de los vertebrados y que están demasiado limitados?
Spinophorosaurus virtual: un nuevo cambio de interpretación
Los estudios llevados a cabo en Spinophorosaurus nigerensis, un saurópodo temprano excepcionalmente completo y bien preservado del período Jurásico de Níger (casi 170 millones de años), han combinado ambas líneas de evidencia, mostrando que lejos de ser incompatibles son perfectamente complementarias. Spinophorosaurus se conoce a partir de tres ejemplares articulados (Remes et al., 2009): un espectacular esqueleto casi completo y articulado de un subadulto de 11 metros de longitud, restos parciales de un adulto de 13 metros de longitud y el cuello y las primeras vértebras de la espalda de un juvenil de 3 metros.
A diferencia de trabajos previos, donde se digitalizaron ilustraciones en dos dimensiones para generar una proyección tridimensional, los huesos de estos tres Spinophorosaurus fueron todos digitalizados de forma individual mediante una técnica llamada fotogrametría digital, con el fin de elaborar un esqueleto virtual completo de Spinophorosaurus. Esta técnica, basada en el mismo principio básico que un estereoscopio, consiste en la toma de muchas fotografías ligeramente superpuestas que recojan toda la superficie de cada uno de los huesos fósiles del esqueleto (Mallison y Wings, 2014). Su procesamiento en un ordenador permite obtener muchísimos puntos homólogos entre todas las fotografías, que se usan para generar una superficie virtual que reproduce el fósil con un margen de error inferior al milímetro –si el escaneo se ha llevado a cabo correctamente–. Repetido este proceso con cada hueso, se obtiene una colección de huesos virtuales listos para ser articulados.
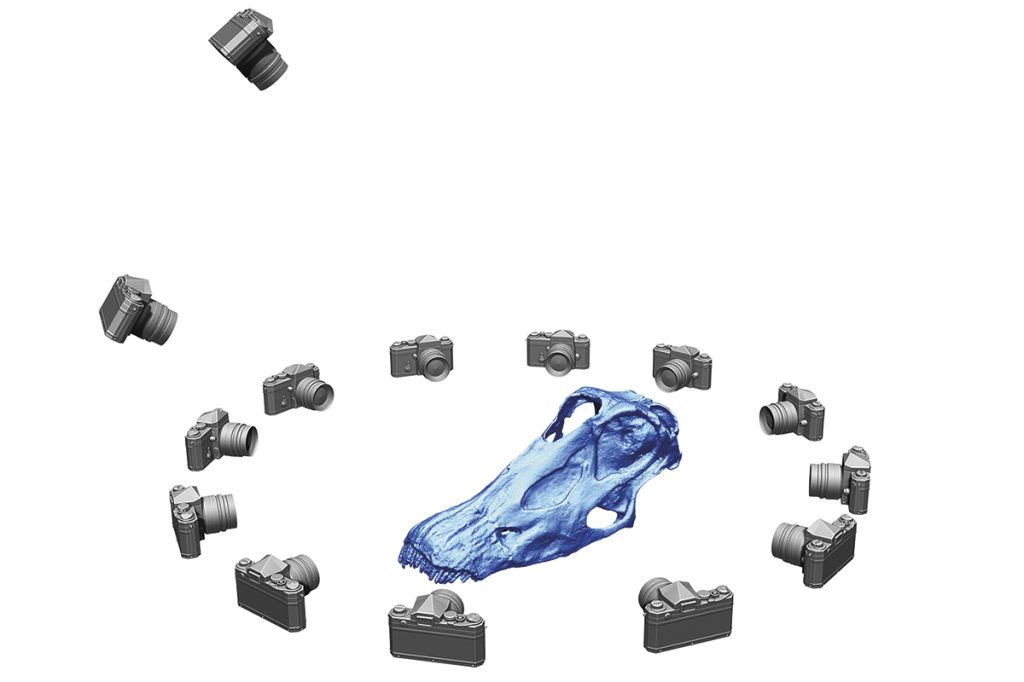
El primer paso para tener una noción de cómo se dispone el esqueleto de un vertebrado es articular los huesos en lo que se denomina postura osteológica neutra, aquella posición que resulta de articular los huesos en el máximo contacto que permiten sus facetas de articulación. Aunque se denomina neutra, esto no implica necesariamente que sea la postura más habitual que adopta un animal en vida (Dzemski y Christian, 2007). Se trata de un punto de partida relativamente objetivo, que permite comparar la disposición del esqueleto entre distintas especies de vertebrados bajo un mismo criterio: el máximo contacto articular. Intentando evitar nociones preconcebidas de cuál pudiera ser esta postura osteológica neutra, la columna vertebral se articula únicamente con una pareja de vértebras visibles a la vez. Manteniendo una vértebra completamente estática, se articula la siguiente con la totalidad de sus facetas de articulación en contacto. Una vez articuladas estas dos primeras vértebras, el software permite volver invisible la primera vértebra e importar una nueva vértebra. Ahora podemos ver solo el segundo y el tercer hueso, y podemos ajustar la posición del tercer hueso para que encaje perfectamente con el segundo hueso. Proceder de esta manera elimina cualquier sesgo que las nociones preconcebidas pudieran introducir en la postura neutra de la columna vertebral. Estas nociones preconcebidas aún hoy en día influyen muchísimo en cómo se articulan los esqueletos de saurópodos en numerosos museos, donde muchas veces se tiene que forzar la desarticulación de la columna vertebral en varios puntos para obtener una postura con la espalda arqueada, por ejemplo.
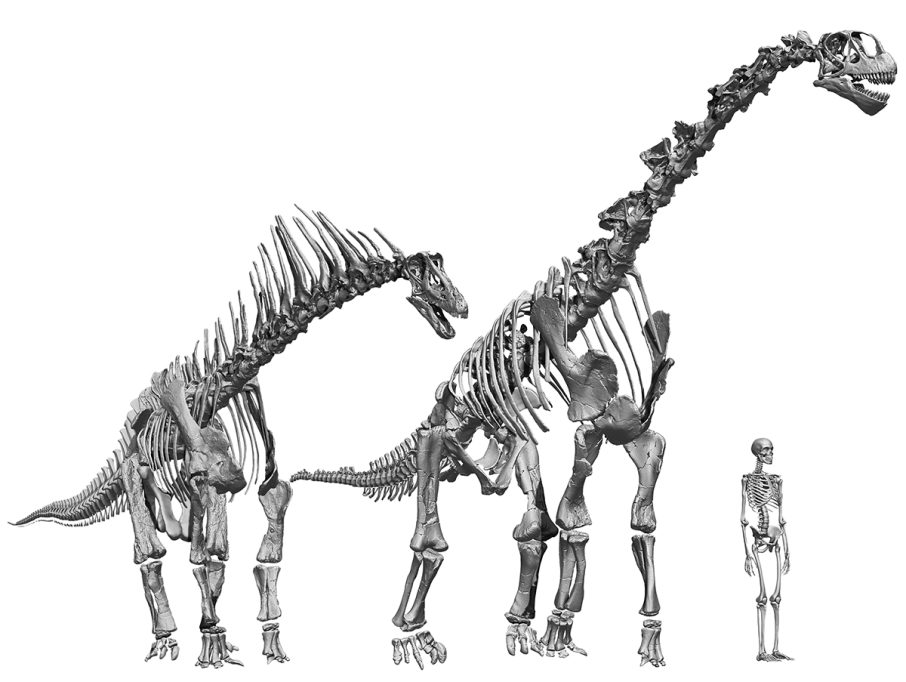
Sorprendentemente, el esqueleto virtual resultante de aplicar este protocolo fue radicalmente diferente de todas las reconstrucciones anteriores de este dinosaurio, que mostraban a Spinophorosaurus con una columna vertebral horizontal. Sin embargo, la nueva reconstrucción mostraba un cuello y una espalda inclinados hacia arriba con una cola horizontal (Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020). A pesar de su elevación, el cuello quedaba alineado prácticamente con las vértebras dorsales, sin cambios en el ángulo de elevación en la base del cuello, como habían demostrado estudios anteriores (Stevens y Parrish, 2005). Pero, ¿cómo es posible que Spinophorosaurus tenga el cuello inclinado hacia arriba? Tenía que haber un punto donde la columna vertebral se elevara, donde estuviera la «clave de bóveda» de los saurópodos. Y encontramos esa piedra angular en las vértebras del sacro, las vértebras fusionadas con los huesos de la cadera.
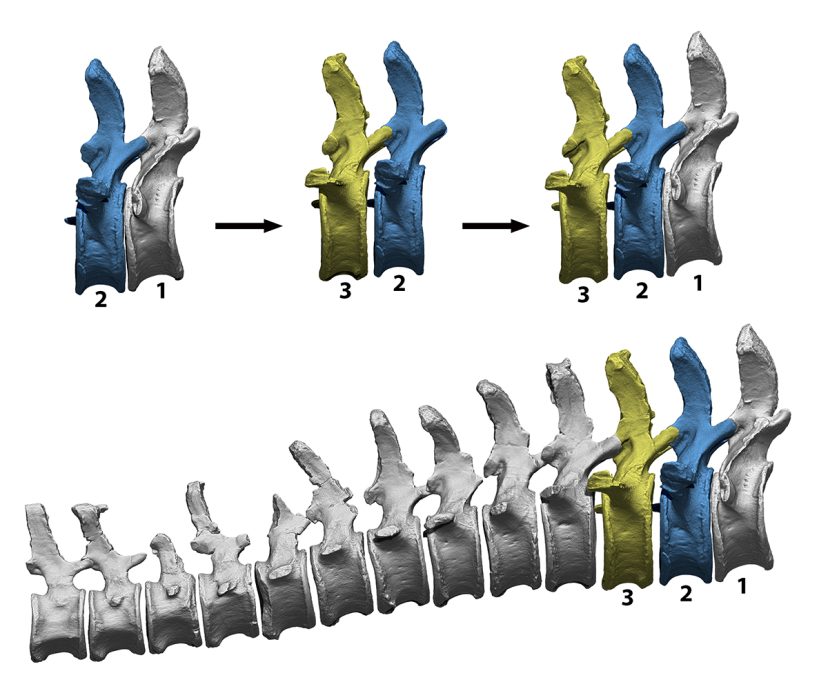
Proceso de articulación en postura neutra de una secuencia de vértebras caudales virtuales del dinosaurio saurópodo Camarasaurus, con el que evitar que nociones preconcebidas tengan un impacto significativo en la curvatura de la columna vertebral. Primero se articula el primer par de vértebras de forma que el solapamiento de sus superficies de articulación es máximo. Después se oculta la primera vértebra caudal y se añade la tercera, estando siempre visibles una pareja de ellas y articulándose mediante el mismo criterio. La curvatura resultante es fruto del máximo contacto articular, lo cual minimiza posibles sesgos de interpretación.
Esto resultó sorprendente, ya que la mayoría de los paleontólogos esperaban que el punto clave fuera donde el cuello se une con la espalda o la base del cuello (Stevens, 2013; Taylor et al., 2009). Sin embargo, como dijo el paleontólogo Henry F. Osborn (1899), igual que en las aves, «la unión sacropélvica es el centro de la energía y del movimiento», es el área más reforzada de la columna vertebral; por tanto, la que podría soportar modificaciones sin comprometer su capacidad para que el animal caminara. El sacro de Spinophorosaurus tiene forma de cuña o piedra angular: es más largo en su parte inferior y más corto en la mitad superior. Por consiguiente, cuando se articula la cola, el torso y el cuello se elevan hacia arriba, lo que permitía que desde esta postura neutra Spinophorosaurus situara su hocico a cinco metros del suelo, más del doble de altura que su hombro.
¿Eran los saurópodos como las jirafas?
Para saber si estas técnicas realmente son capaces de desentrañar datos precisos de la paleobiología de los dinosaurios saurópodos, la aproximación más inmediata es evaluar si es posible replicar con ellas las capacidades biológicas de vertebrados actuales. La jirafa es el único vertebrado actual que puede utilizarse como modelo para los cuellos de los saurópodos: son animales cuadrúpedos de gran tamaño que superan la tonelada de peso y con un largo cuello, fruto de la elongación de sus vértebras cervicales y la «cervicalización» de su primera vértebra torácica. Aplicando sobre esqueletos de jirafas actuales las mismas técnicas descritas para los saurópodos, las reconstrucciones esqueléticas obtenidas coinciden perfectamente con las posturas habitualmente adoptadas por las jirafas al caminar (Vidal, Mocho, Páramo et al., 2020).
En la naturaleza, las jirafas se alimentan habitualmente con el cuello situado por encima de sus hombros, pero ocasionalmente los machos se alimentan también, con el cuello completamente vertical, de los brotes más altos que puedan alcanzar y las hembras se alimentan pastando a ras de suelo. Todo este rango de posiciones de los cuellos de las jirafas se puede obtener con sus esqueletos virtuales sin necesidad de desarticular ninguna vértebra. Es más, incluso la postura de máxima ventriflexión del cuello de las jirafas virtuales no permite alcanzar el suelo sin flexionar los codos o espatarrar los hombros, exactamente igual que ocurre con las jirafas vivas. Se puede afirmar que todo el repertorio de posiciones del cuello de las jirafas se puede replicar virtualmente con los mismos parámetros utilizados para los saurópodos.
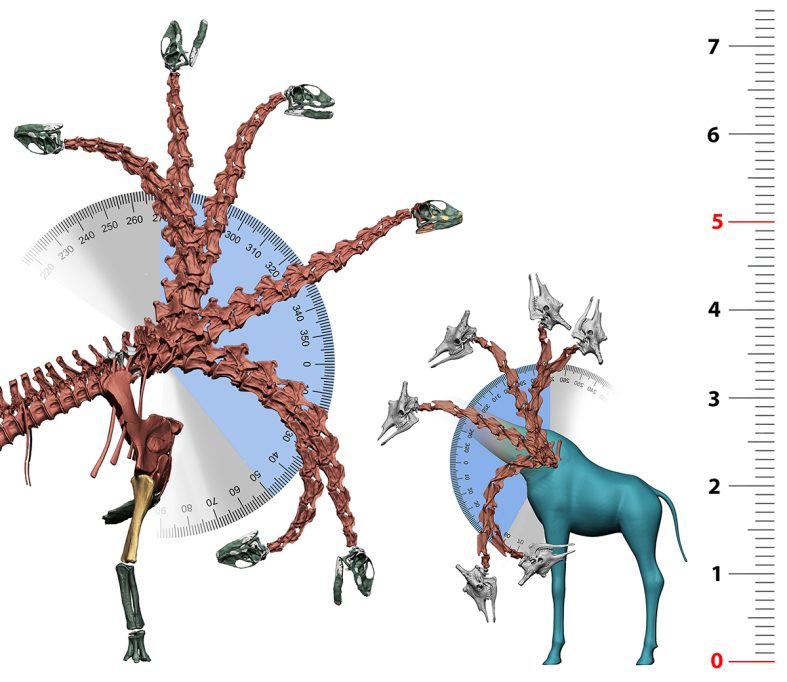
¿Qué implica esto para los dinosaurios saurópodos? Esta es una cuestión más compleja que afirmar que, si un organismo extinto podía adoptar físicamente una postura, esta era adoptada habitualmente y formaba parte de su comportamiento. Los seres humanos somos capaces de caminar sobre la punta de los dedos de los pies al bailar ballet y los elefantes son capaces de sostenerse a dos patas sobre sus extremidades delanteras. Sin embargo, esto no implica que bailar ballet o sostenerse con las extremidades delanteras sean comportamientos habituales de ninguna de esas dos especies (Mallison, 2010; Sander et al., 2010). Sin embargo, esa capacidad existe. Las técnicas de paleontología virtual permiten desvelar qué capacidades físicas tenían los vertebrados extintos y qué posturas o comportamientos eran físicamente imposibles y, por lo tanto, deberían descartarse a la hora de interpretar la ecomorfología de estos organismos (Mallison, 2010). En el caso de Spinophorosaurus nigerensis se puede afirmar que su esqueleto le permitía adoptar posiciones que le hubieran permitido alimentarse del mismo modo que las jirafas actuales: desde copas de árboles a casi 8 metros de altura hasta vegetación a nivel del suelo. Es uno de los primeros saurópodos de los que tenemos evidencia de estas capacidades. Otros saurópodos, como los diplodocoideos Apatosaurus, Diplodocus o Amargasaurus, no tenían esqueletos que les confirieran las mismas capacidades de movimiento y estaban más restringidos a la hora de situar el cuello verticalmente, lo cual es evidencia de que ocuparían probablemente nichos ecológicos diferentes (Christian et al., 2005; Stevens y Parrish, 1999, 2005). Por lo tanto, los modelos virtuales sí que permiten reproducir las capacidades funcionales de los vertebrados actuales y consecuentemente también inferir las de los vertebrados extintos. Pese a ello, no es posible saber si todas estas capacidades formaban parte de su comportamiento o, si así era, si eran capacidades usadas habitualmente o de forma esporádica. Sin embargo, es posible utilizar estas técnicas para refutar comportamientos, siempre y cuando estos sean incompatibles con las capacidades de un organismo, y la paleontología virtual es capaz de mostrarlo.
La cópula en los dinosaurios saurópodos
Un misterio difícil de conocer es la interpretación correcta de cómo se reproducían los dinosaurios no avianos. Es seguro que los dinosaurios, incluso los saurópodos más grandes, podían realizar la cópula, puesto que todos ellos tuvieron ascendencia y descendencia, pero su gigantesco tamaño nos hace preguntarnos cómo era exactamente. Para hacerse una idea de cómo podría ser esta cópula, lo primero es determinar cómo la realizan los parientes vivos más cercanos a los dinosaurios saurópodos, es decir, los cocodrilos y las aves. La cópula en cocodrilos es muy similar en todas las especies actuales y es la misma que en muchos reptiles: el macho se sitúa junto a la hembra y coloca su extremidad posterior sobre la cadera de la hembra, una posición llamada en inglés leg over back (literalmente, ‘pierna sobre la espalda’). Esta posición facilita que el «pene» del macho pueda introducirse en la cloaca de la hembra, puesto que el apéndice caudal de los reptiles suele dificultar la cópula (Isles, 2009). Las aves redujeron muchísimo su apéndice caudal durante su evolución, y en la mayoría de aves actuales está formado por unas pocas vértebras con movilidad, de modo que no supone un impedimento para la cópula. Por este motivo, las aves no realizan la postura de leg over back como sí que hacen los cocodrilos. Las aves más primitivas tienen un «pene» (de función análoga al de los mamíferos, aunque de origen evolutivo diferente), pero los paseriformes y galliformes tienden a reducirlo o perderlo por completo (Isles, 2009).
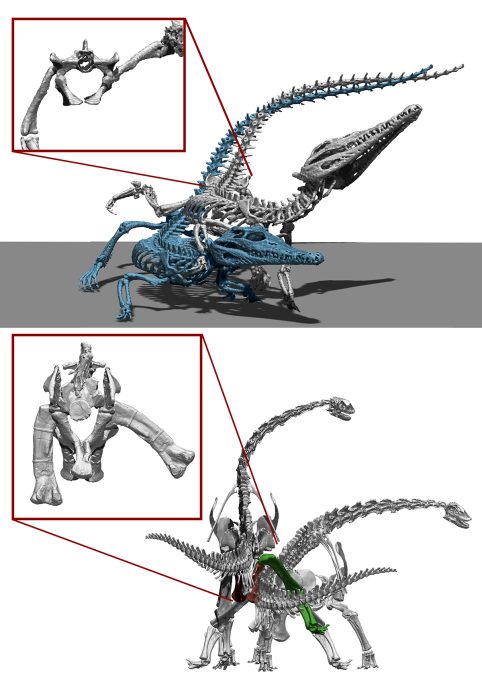
El hecho de que los cocodrilos tengan un plan corporal más similar al de los saurópodos (tienen un apéndice caudal bien desarrollado y son cuadrúpedos) sugiere que tal vez estos últimos adoptaran también el leg over back como estrategia para facilitar la cópula (Hallet y Wedel 2016; Isles, 2009). Sin embargo, el esqueleto de los saurópodos simplemente no permitiría esta posición. A diferencia de los cocodrilos, cuyo fémur tiene una cabeza redondeada que permite mucha movilidad en el acetábulo de la cadera, los saurópodos tienen fémures con una cabeza en forma de ele invertida. Esto causa que cualquier intento de abducción pronto se vea limitado al colisionar con las paredes del acetábulo, reduciendo mucho su movimiento en ese plano. La limitada capacidad de los saurópodos para elevar su extremidad posterior implica que no existiría suficiente espacio entre la cadera y la extremidad posterior para acomodar a la hembra, como sí que sucede en los cocodrilos, a menos que esta fuera mucho más pequeña que el macho. Existen pocos estudios sobre el dimorfismo sexual en dinosaurios saurópodos, pero la poca evidencia existente indica que no había un dimorfismo muy acusado en el tamaño entre ambos sexos (Ikejiri, 2004). Por consiguiente, se puede asegurar que los saurópodos no podían adoptar la misma posición de cópula que los cocodrilos y una gran mayoría de reptiles actuales, y que sus estrategias de cópula debían de ser distintas. Otras posibilidades que hubieran podido adoptar los saurópodos son una monta similar a la que realizan la mayoría de los mamíferos cuadrúpedos, en la que el macho se situaría sobre sus extremidades traseras y se apoyaría con las delanteras sobre la hembra, que podría situarse de pie o recostada. No existe ningún impedimento en el rango de movimiento osteológico de los saurópodos que impidiera llevar a cabo estas posiciones. Sin embargo, tampoco hay ningún impedimento osteológico a que pudieran realizar una cópula de espaldas, de manera similar a cómo la realizan muchos artrópodos y algunos cánidos. En definitiva, no podemos saber cómo se apareaban los saurópodos, puesto que no podemos refutar todas las posibilidades. Lo que sí que podemos afirmar con absoluta certeza es que no lo hacían del mismo modo que los cocodrilos actuales.
Conclusión: una herramienta adicional y complementaria
Una vez vistos estos ejemplos, es evidente que las tecnologías virtuales no son una novedad que ha llegado para sustituir a la paleontología y la anatomía comparadas tradicionales por haberse quedado obsoletas, sino que son una herramienta adicional y complementaria para poder evaluar de forma científica algunas hipótesis que antes resultaban muy difíciles o incluso imposibles de ser testadas. Hemos visto cómo la paleontología virtual muestra que la hipótesis tradicional de los saurópodos como análogos de las jirafas era totalmente matizable, y que algunos saurópodos tenían capacidades para alimentarse igual que las jirafas, mientras que otros tenían capacidades de movimiento más limitadas en sus cuellos, lo cual les condicionaba a alimentarse de vegetación de bajo porte o a ras de suelo. Esto sugiere un escenario más complejo a los considerados previamente, donde no se trata de si todos los saurópodos podían o no podían alimentarse como las jirafas, sino de cuáles podían y cuáles no podían hacerlo.
Del mismo modo, hasta hace poco, cómo se apareaban los saurópodos era una cuestión cercana a la especulación. Sin embargo, gracias a la paleontología virtual se ha podido refutar que los saurópodos se apareasen como lo hacen los cocodrilos actuales. La paleontología virtual no ha dejado de requerir de observaciones en animales actuales vivos o de contar con ejemplares fósiles excepcionalmente completos para poder evaluar estas hipótesis de forma contundente. Ha supuesto a la vez una revolución en cómo estudiamos los fósiles de dinosaurios, pero también un recordatorio de la importancia de la confluencia de múltiples líneas de evidencia como mejor forma de avanzar el conocimiento científico.
Bakker, R. T. (1971). Ecology of the brontosaurs. Nature, 229, 172–174. https://doi.org/10.1038/229172a0
Dzemski, G., & Christian, A. (2007). Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length. Journal of Morphology, 268, 701–714. https://doi.org/10.1002/jmor.10542
Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs. Johns Hopkins University Press.
Ikejiri, T. (2004). Anatomy of Camarasaurus lentus (Dinosauria: Sauropoda) from the Morrison formation (Late Jurassic), Thermopolis, Central Wyoming, with determination and interpretation of ontogenetic, sexual dimorphic and individual variation in the genus. Fort Hays State University.
Isles, T. E. (2009). The socio-sexual behaviour of extant archosaurs: Implications for understanding dinosaur behaviour. Historical Biology, 21(3–4), 139–214. https://doi.org/10.1080/08912960903450505
Mallison, H. (2010). CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915. Swiss Journal of Geosciences, 103(2), 211–233. https://doi.org/10.1007/s00015-010-0024-2
Mallison, H., & Wings, O. (2014). Photogrammetry in paleontology–A practical guide. Journal of Paleontological Techniques, 12, 1–31.
Osborn, H. F. (1899). A skeleton of Diplodocus, recently mounted in the American Museum. Science, 10(259), 870–874. https://doi.org/10.1126/science.10.259.870
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Ferrer, J. M. M., Ide, O. A., & Maga, A. (2009). A new basal sauropod dinosaur from the middle Jurassic of Niger and the early evolution of Sauropoda. PLOS ONE, 4(9), e6924. https://doi.org/10.1371/journal.pone.0006924
Sander, P. M., Christian, A., Clauss, M., Fechner, R., Gee, C. T., Griebler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S. F., Preuschoft, H., Rauhut, O. W. M., Remes, K., Tutken, T., Wings, O., & Witzel, U. (2010). Biology of the sauropod dinosaurs: The evolution of gigantism. Biological Reviews, 86, 117–155. https://doi.org/10.1111/j.1469-185X.2010.00137.x
Sereno, P. C., Wilson, J. A., Witmer, L. M., Whitlock, J. A., Maga, A., Ide, O., & Rowe, T. A. (2007). Structural extremes in a cretaceous dinosaur. PLOS ONE, 2(11), 1–9. https://doi.org/10.1371/journal.pone.0001230
Stevens, K. A. (2013). The articulation of sauropod necks: Methodology and mythology. PLOS ONE, 8(10), 1–27. https://doi.org/10.1371/journal.pone.0078572
Stevens, K. A., & Parrish, M. J. (1999). Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science, 284, 798–800. https://doi.org/10.1126/science.284.5415.798
Stevens, K. A., & Parrish, M. J. (2005). Neck posture, dentition, and feeding strategies in Jurassic sauropod dinosaurs. En V. Tidwell & K. Carpenter (Eds.), Thunder-lizards: The sauropodomorph dinosaurs (pp. 212–232). Indiana University Press.
Sutton, M. D., Rahman, I. A., & Garwood, R. J. (2014). Techniques for virtual palaeontology. Wiley. https://doi.org/10.1002/9781118591192
Taylor, M. P. (2010). Sauropod dinosaur research: A historical review. Geological Society London Special Publications, 343(1), 361–386. https://doi.org/10.1144/SP343.22
Taylor, M. P., Wedel, M. J., & Naish, D. (2009). Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica, 54(2), 213–220. https://doi.org/10.4202/app.2009.0007
Vidal, D., Mocho, P., Aberasturi, A., Sanz, J. L., & Ortega, F. (2020). High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs. Scientific Reports, 10(1), 6638. https://doi.org/10.1038/s41598-020-63439-0
Vidal, D., Mocho, P., Páramo, A., Sanz, J. L., & Ortega, F. (2020). Ontogenetic similarities between giraffe and sauropod neck osteological mobility. PLOS ONE, 15(1). https://doi.org/10.1371/journal.pone.0227537




