

El mapa de la biodiversidad
De la escala local a la global

La riqueza de especies no es homogénea en el espacio y normalmente presenta diferencias cuando comparamos diferentes lugares. Estas diferencias suelen responder a gradientes de uno o varios factores que crean patrones espaciales de biodiversidad y dependen de la escala. A escala local, los patrones de diversidad dependen del tamaño del hábitat (la relación especies-área), de la productividad, la hostilidad del entorno, la frecuencia e intensidad de las perturbaciones o la reserva regional de especies. La diversidad regional se puede ver afectada por la heterogeneidad ambiental (la cual aumenta la disimilitud), aunque también puede actuar a escalas espaciales mayores o menores, y la conectividad entre diferentes hábitats. Por último, a escala global, los patrones de diversidad se corresponden con la latitud, la altitud o la profundidad, aunque estos factores representan una o más variables ambientales (productividad, área, aislamiento u hostilidad).
Palabras clave: riqueza de especies, relación especies-área, productividad, gradiente latitudinal de biodiversidad.
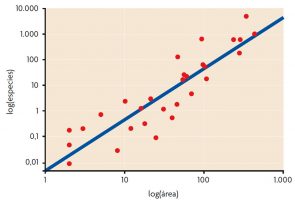
Figura 1. Relación especies-área de las plantas vasculares del archipiélago de Galápagos: un incremento del área de muestra resulta en un incremento del número de especies encontradas. Elaborada a partir de Connor y McCoy, 2000 / Encyclopedia of Biodiversity
¿Por qué algunos lugares son más diversos que otros?
Un rápido vistazo a nuestro entorno cercano debería ser suficiente para que nos diésemos cuenta de que la distribución de especies no es homogénea en el espacio y que algunos lugares contienen más especies que otros con la misma superficie. De hecho, las especies presentes en un determinado lugar son el resultado de diversos procesos que operan a diferentes escalas, como las barreras geográficas, las limitaciones ambientales o las interacciones bióticas que determinan la composición de las comunidades ecológicas. Todos estos procesos definen la composición de especies en cada comunidad: el número y la identidad de especies (o la biodiversidad) en una zona determinada. ¿Qué factores específicos influyen en esta distribución heterogénea de la diversidad en el espacio? Esta pregunta ha despertado el interés de los naturalistas desde que comenzaron a explorar el mundo. Con el paso del tiempo observaron que la biodiversidad de muchos grupos taxonómicos seguía patrones espaciales predecibles, por lo que la distribución de la biodiversidad en el espacio se podía explicar gracias a ciertos factores, y los patrones observados se repetían en muchas regiones del mundo.
«Los naturalistas observaron que la biodiversidad de muchos grupos taxonómicos seguía patrones espaciales predecibles»
Uno de los primeros naturalistas que describió la relación entre los organismos y sus entornos fue Alexander von Humboldt (1769-1859), quien se centró principalmente en el efecto de factores geográficos como el clima sobre diferentes taxones. El trabajo de Humboldt en su expedición científica a América inspiró profundamente a otros naturalistas como Charles Darwin (1809-1882) o Alfred Russel Wallace (1823-1913) a llevar a cabo sus propias exploraciones del mundo. Todas estas expediciones condujeron a la posterior búsqueda de patrones de distribución de especies y de diversidad a gran escala. Desde entonces, se han propuesto muchas hipótesis para contestar esa pregunta, pero los factores específicos que subyacen a los gradientes de diversidad siguen siendo controvertidos. Además, aunque algunos de los patrones pueden actuar a diferentes escalas, la mayoría dependen de la escala en sí; por tanto, en función de la escala espacial que se estudie, los factores relacionados con los gradientes de diversidad cambiarán.
«Probablemente los patrones de biodiversidad a escala local fueron los primeros en ser descritos, como resultado de observaciones de campo básicas y sencillas»
Es importante subrayar que muchos estudios que analizan patrones espaciales de biodiversidad utilizan el número de especies (la riqueza de especies) como variable de respuesta. Recientemente, los biólogos han empezado a centrarse en otros aspectos de la biodiversidad como la diversidad genética o funcional. Sin embargo, por una cuestión de simplicidad, centraremos este repaso solo en la diversidad de especies. Por lo tanto, de aquí en adelante utilizaremos de forma indistinta los términos biodiversidad y riqueza de especies. A continuación discutiremos en más detalle los patrones de biodiversidad a diferentes escalas espaciales (resumidos en la Tabla 1).

Figura 2. Uno de los patrones de la relación entre especies y área indica que las islas más grandes de un archipiélago albergan un mayor número de especies. Esta es una de las bases de la teoría de biogeografía de islas de MacArthur y Wilson. En la imagen, foto satélite del archipiélago de Hawái (EE UU). / Jacques Descloitres, MODIS Land Rapid Response Team, NASA GSFC Figura 3. Existen factores que determinan los patrones de biodiversidad y limitan el número de especies, por ejemplo, la hostilidad del ambiente, como ocurre con temperaturas extremas. En la imagen, el valle desértico conocido como Wadi Rum, en Jordania. / Martino Pietropoli
Patrones locales
Probablemente los patrones de biodiversidad a escala local fueron los primeros en ser descritos, como resultado de observaciones de campo básicas y sencillas. Uno de los primeros patrones en ser observado fue la relación entre número de especies y área: cuanto mayor es el área muestreada, mayor es el número de especies que alberga (Figura 1). Este patrón se ha observado en todo el mundo, tanto en entornos terrestres como en marinos. La primera descripción de la relación especies-área se realizó en el siglo XIX, cuando H. C. Watson observó en un condado de Inglaterra que el número de especies de plantas en una muestra se doblaba al incrementar diez veces el área estudiada (Connor y McCoy, 2000). Después, Arrhenius cuantificó esta relación por primera vez en 1921 mediante la función S = cAz (donde S es el número de especies, A es el área y c y z son constantes). Las constantes c y z sirven para establecer comparaciones entre diferentes áreas de estudio.
«Uno de los primeros patrones en ser observados fue la relación entre número de especies y área: cuanto mayor es el área muestreada, mayor es el número de especies que alberga»
Rosenzweig (1997) considera que en realidad las curvas especies-área se podrían dividir en cuatro patrones distintos. Los dos primeros dependen del tamaño del área muestreada: pequeña o grande. Un tercero considera la macroescala (provincias biogeográficas), en las que el incremento de especies no se relaciona con la inmigración de especies desde otras áreas, sino con los procesos de especiación, que actúan a una escala temporal más lenta. Y el cuarto patrón es la base de la teoría de biogeografía de islas de MacArthur y Wilson, que indica que las islas más grandes de un archipiélago albergan un mayor número de especies (Figura 2). Se utilizan dos argumentos principales para explicar la relación entre número de especies y área: primero, que las áreas más grandes pueden mantener poblaciones más grandes y, por lo tanto, las especies tienen menos probabilidades de extinguirse; y segundo, que los hábitats con superficies más grandes son más heterogéneos (explicado más adelante a escala regional).
| Escala | Factor |
| Local | Área |
| Productividad del ecosistema | |
| Hostilidad del entorno | |
| Nivel de perturbación | |
| Reserva regional de especies | |
| Regional | Heterogeneidad ambiental |
| Conectividad | |
| Global | Latitud |
| Altitud (en montañas) | |
| Profundidad (en ecosistemas marinos) |
Tabla 1. Resumen de los diferentes factores que crean patrones espaciales de diversidad y las escalas en las que operan: local, regional y global. En algunos casos, el factor no es la causa que subyace al gradiente de diversidad, sino que es una suma de diferentes variables la que crea el patrón (por ejemplo la latitud; véase el texto).
Otro patrón que se ha estudiado bastante es la relación entre riqueza de especies y productividad en un ecosistema. La productividad es el ritmo al que se produce biomasa en un área determinada. Es una medida del aporte energético a un ecosistema (normalmente estimado mediante la precipitación, evapotranspiración o aportación de nutrientes). Originalmente se pensaba que los recursos tendían a ser ilimitados en los niveles más altos de productividad, lo que permitía la presencia de un número mayor de especies. Este patrón se observa principalmente a escala global, donde las regiones biogeográficas con más aporte energético generalmente contienen más especies (véase el apartado de patrones globales). A escalas locales se han observado diferentes tipos de relaciones: positiva, negativa, en forma de U y en forma de campana (Mittelbach et al., 2001). Esta última la encontramos cuando la biodiversidad es mayor con niveles intermedios de productividad, y es lo suficientemente frecuente en la naturaleza como para haber motivado numerosos estudios que intentan explicar sus causas. En la primera parte del gradiente de productividad (productividad baja o media) la diversidad de especies aumenta conforme aumenta la disponibilidad de recursos. Sin embargo, la disminución en la riqueza de especies que se produce a partir de cierto nivel de productividad (la paradoja del enriquecimiento) no está tan clara. Se han sugerido diversas hipótesis, como el aumento de la exclusión competitiva cuando existen niveles altos de productividad o el cambio en el recurso limitante para las plantas, que pasa de ser los nutrientes a ser la luz (Tilman y Pacala, 1993). No obstante, ninguna hipótesis explica la variación en forma y magnitud de la relación entre productividad y diversidad, y factores como la escala espacial, los taxones estudiados, el tipo de hábitat (terrestre o acuático) o la intensidad de la depredación (coexistencia mediada por el explotador) pueden ser relevantes para explicar esta disparidad.
Aunque tanto el área como la productividad favorecen la coexistencia de numerosas especies, otros factores determinan los patrones de biodiversidad limitándolas en número. Por ejemplo, la hostilidad del entorno (hábitats muy alcalinos o ácidos, o con temperaturas extremas) selecciona las especies que pueden sobrevivir en ellos, organismos con adaptaciones muy específicas (Figura 3). Sin embargo, para complicar más las cosas, puede que estos hábitats también compartan características comunes que podrían reducir la riqueza de especies, como un área pequeña o cierto grado de aislamiento (por ejemplo, aguas termales o cumbres montañosas). Esto dificulta la posibilidad de separar el efecto de todos estos factores en la diversidad local. Otra hipótesis propuesta para explicar los gradientes de diversidad es la hipótesis de la perturbación intermedia (Connell, 1978), que sugiere que los hábitats con niveles altos y bajos de perturbación contienen pocas especies. Por lo tanto, la máxima diversidad se encuentra en los niveles intermedios de perturbación (Connell, 1978), porque estos impiden la dominancia de buenos colonizadores (abundantes con niveles altos de perturbación) y competidores (con niveles bajos). Aunque la primera demostración empírica se realizó en rocas intermareales sometidas a diferentes grados de intensidad de tormentas (Sousa, 1979), los estudios realizados con otros sistemas no han logrado encontrar pruebas que apoyen la hipótesis de la perturbación intermedia.
«Las primeras pruebas de los gradientes de biodiversidad a gran escala nos las proporcionaron las primeras expediciones europeas al nuevo mundo después del siglo XVIII»
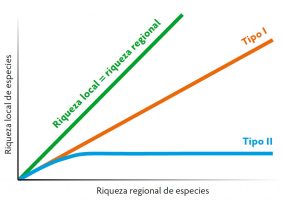
Finalmente, el tamaño de la reserva regional de especies es relevante para la diversidad local. ¿Qué ocurre en comunidades locales en las que existe un gradiente en la riqueza regional de especies? En teoría, se podrían observar dos respuestas distintas (Figura 4). La primera la encontramos cuando la composición de las comunidades locales depende en gran medida de la reserva regional de especies (especies que llegan y colonizan las comunidades locales). En este caso, encontraremos una relación lineal positiva: conforme aumente la diversidad regional, también lo hará la diversidad local (curva de tipo I). La segunda se producirá cuando las comunidades locales presenten algunas características que limiten la entrada indiscriminada de especies de la región (por ejemplo, competencia o depredación). En este caso, la riqueza local se saturará con la riqueza regional porque el número de espacios ecológicos en los hábitats locales es limitado (curva de tipo II). Los estudios empíricos han concluido que la respuesta de tipo I es la más común en la naturaleza.

Patrones regionales
Se considera que una región incluye un gran número de hábitats y comunidades, y a menudo se hace referencia a ella como el área desde la cual las especies colonizan comunidades locales. Por tanto, uno de los principales factores que crean un gradiente de diversidad a escala regional es la heterogeneidad ambiental: si el entorno de la región es homogéneo, sus comunidades probablemente contengan las mismas especies, lo que resulta en una riqueza regional de especies baja.1 Por el contrario, una variación amplia de las condiciones ambientales en diferentes hábitats de una región permitirá la presencia de comunidades diversas (muy disimilares)2 y una riqueza regional de especies alta (Figura 5). La relación positiva entre heterogeneidad ambiental y riqueza de especies se ha demostrado de manera empírica en diferentes taxones, ecosistemas y escalas espaciales (Stein, Gerstner y Kreft, 2014). Además de la variedad de hábitats en una región, este patrón también opera a escalas espaciales menores (heterogeneidad de microhábitats a escala local) y mayores (gradientes de hábitats a escala global).
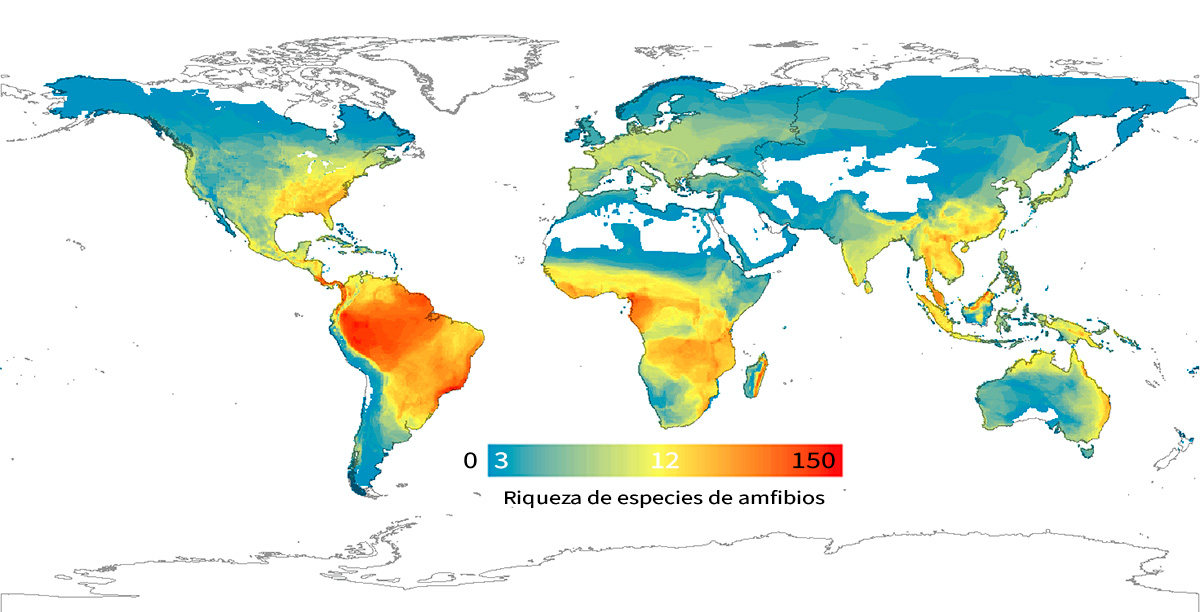
Figura 6. Gradiente latitudinal de diversidad en anfibios, que incluye 6.117 especies en celdas de 5 grados de latitud. El número de especies es mayor en los trópicos y disminuye gradualmente hacia los polos. El gradiente de colores indica una variación que va de 3 especies por celda (azul), a 12 (amarillo) y a 150 (rojo) aproximadamente. Fuente: Pyron y Wiens, 2013. Publicado por la Royal Society. Todos los derechos reservados.
En las últimas décadas, con el creciente interés en las metacomunidades (conjunto de comunidades locales conectadas por dispersión), el flujo de individuos entre parcelas locales se ha empezado a tener en cuenta. Uno de los factores importantes que afectan a este flujo es el grado de conectividad de las parcelas locales, determinado por la localización de cada parcela en el paisaje (Borthagaray, Pinelli, Berazategui, Rodríguez-Tricot y Arim, 2015). Así, dependiendo de la configuración de la red de la metacomunidad, así como de la capacidad de dispersión de las especies y el tamaño de cada una de ellas, la riqueza de especies local y regional puede variar. Por ejemplo, en el caso de organismos con tasas de dispersión bajas, cuando las parcelas están aisladas, la diversidad local sería baja, pero la regional sería alta. Por el contrario, cuando los organismos se dispersan fácilmente y las parcelas están muy interconectadas, la diversidad local sería alta y la regional baja.

Figura 7. Ilustración del «efecto de dominio medio» con lápices para mostrar una de las posibles explicaciones del gradiente latitudinal de diversidad (la disminución de diversidad desde el ecuador a los polos o en cualquier área con unos límites claros): por puro azar, el centro de una caja llena de lápices de diferentes tamaños contendrá más lápices (o más partes) que los extremos. Elaborada a partir de Mittelbach, 2012 / Community Ecology
Patrones globales
Como ya se indicó anteriormente, las primeras evidencias de gradientes de biodiversidad a gran escala nos las proporcionaron las primeras expediciones europeas al Nuevo Mundo después del siglo XVIII. En estas exploraciones, se envió a los naturalistas a describir las especies exóticas del lugar para conseguir nueva información acerca de la misteriosa y exótica naturaleza de este nuevo continente. Muchos de ellos quedaron fascinados por la gran diversidad de especies (y sus formas, colores y comportamientos) de los trópicos en comparación con las bien conocidas regiones europeas. Estas observaciones condujeron a la descripción del patrón de biodiversidad a gran escala más famoso: el gradiente latitudinal de diversidad, caracterizado por una disminución en la riqueza de especies desde el ecuador hacia latitudes mayores (al norte o al sur) (Figura 6). Esta disminución en la diversidad es ligeramente asimétrica entre los dos hemisferios, con una curva más pronunciada en el hemisferio norte. El gradiente latitudinal de diversidad se ha documentado en una variedad de taxones animales y vegetales de diferentes ecosistemas (marinos y terrestres; Hillebrand, 2004). Sin embargo, existen algunas excepciones. Por ejemplo, los macrófitos acuáticos representan uno de los pocos grupos taxonómicos que muestran un gradiente latitudinal de diversidad inverso, con comunidades más ricas en latitudes más altas.
Además, este gradiente no se observa solo en especies actuales, sino también en el registro fósil (se ha observado sobre todo en algunos taxones de invertebrados marinos, como los braquiópodos o los foraminíferos). No obstante, la latitud en sí misma no es la causa subyacente de este gradiente de biodiversidad. Esta correlación podría explicarse con muchos factores ambientales. Hasta ahora se han sugerido muchos mecanismos diferentes y, probablemente, la combinación de algunos de ellos afecte a la variación en la diversidad a lo largo del gradiente latitudinal. Mittelbach (2012) resume todas las posibles hipótesis en cuatro grupos: ecológicas, históricas, evolutivas y el modelo nulo. Las hipótesis ecológicas se basan principalmente en la relación entre productividad (el aporte energético y la disponibilidad de recursos) y una mayor abundancia de individuos, lo que reduce el riesgo de extinción. Las hipótesis históricas defienden que los trópicos son más antiguos y que tienen una mayor extensión geográfica (en el pasado y en la actualidad) y unas condiciones climáticas más estables en el tiempo, por lo que han tenido más tiempo para la diversificación. Las hipótesis evolutivas se centran en las tasas más altas de diversificación en los trópicos. Y, finalmente, el modelo nulo se basa en el «efecto de dominio medio»: la idea de que si los rangos de distribución de especies se localizan al azar a lo largo del gradiente latitudinal es más probable que un número mayor de distribuciones de especies se solapen en el medio (en el ecuador). Podemos visualizarlo fácilmente con una caja llena de lápices de diferentes tamaños en la que, por azar, la mayoría de lápices estarán en el centro de la caja (Figura 7).
Aunque el gradiente latitudinal de diversidad es el patrón más apasionante de la ecología, se han descrito otros patrones globales, como los gradientes de elevación en sistemas montañosos y los gradientes de profundidad en entornos marinos. Como la latitud, estas dos variables no son la causa que subyace a los patrones de riqueza de especies. En estos casos, las condiciones ambientales, más hostiles conforme se avanza en el gradiente de altitud y profundidad, así como la baja productividad, el aislamiento o las áreas reducidas (por ejemplo, los picos más altos), pueden contribuir a la disminución en la riqueza de especies.
¿Qué podemos aprender de los patrones espaciales de biodiversidad?
Hemos presentado los patrones de biodiversidad más estudiados a diferentes escalas espaciales. Algunos de ellos presentan una relación clara y generalizada con la diversidad, mientras que en otros casos la respuesta es cuestionable y necesita ser investigada con mayor profundidad. Sin embargo, debemos subrayar que la mayoría de estos patrones se han establecido con importantes lagunas de conocimiento en cuanto a la taxonomía y distribución de especies, especialmente a escala global (hablamos de los conocidos como «déficit de Linneo» y «déficit de Wallace», respectivamente; Hortal et al., 2015). Estas lagunas de conocimiento reflejan no solo las diferencias en los esfuerzos realizados para investigar en diferentes partes de todo el mundo, sino también la desigualdad en los taxones estudiados. Por ejemplo, los conjuntos de datos sobre especies de vertebrados (aves o mamíferos) y algunos grupos de plantas (árboles) son mucho más completos que los de invertebrados u otros taxones más pequeños. De igual forma, los hábitats acuáticos se han estudiado muy poco, pese a su alta y desproporcionada contribución a la diversidad global, y todavía hay cierta incertidumbre sobre si la biodiversidad en los entornos acuáticos sigue los mismos patrones que en los hábitats terrestres (Siqueira, Bini, Thomaz y Fontaneto, 2015). Eliminar estas lagunas ayudará a esclarecer la ubicuidad de los patrones espaciales y sus excepciones (si las hubiera).
Hemos presentado los patrones de biodiversidad más estudiados a diferentes escalas espaciales. Algunos de ellos presentan una relación clara y generalizada con la diversidad, mientras que en otros casos la respuesta es cuestionable y necesita ser investigada con mayor profundidad. Sin embargo, debemos subrayar que la mayoría de estos patrones se han establecido con importantes lagunas de conocimiento en cuanto a la taxonomía y distribución de especies, especialmente a escala global (hablamos de los conocidos como «déficit de Linneo» y «déficit de Wallace», respectivamente; Hortal et al., 2015). Estas lagunas de conocimiento reflejan no solo las diferencias en los esfuerzos realizados para investigar en diferentes partes de todo el mundo, sino también la desigualdad en los taxones estudiados. Por ejemplo, los conjuntos de datos sobre especies de vertebrados (aves o mamíferos) y algunos grupos de plantas (árboles) son mucho más completos que los de invertebrados u otros taxones más pequeños. De igual forma, los hábitats acuáticos se han estudiado muy poco, pese a su alta y desproporcionada contribución a la diversidad global, y todavía hay cierta incertidumbre sobre si la biodiversidad en los entornos acuáticos sigue los mismos patrones que en los hábitats terrestres (Siqueira, Bini, Thomaz y Fontaneto, 2015). Eliminar estas lagunas ayudará a esclarecer la ubicuidad de los patrones espaciales y sus excepciones (si las hubiera).
1. El número de especies que se encuentra en una región se conoce como diversidad gamma. (Volver)
2. Se conoce como diversidad beta la similitud o disimilitud entre comunidades locales. Mide la diferencia en la composición de especies entre dos o más hábitats de una región. (Volver)
Referències
Borthagaray, A. I., Pinelli, V., Berazategui, M., Rodríguez-Tricot, L., & Arim, M. (2015). Effects of metacommunity networks on local community structures: From theoretical predictions to empirical evaluations. En A. Belgrano, G. Woodward, & U. Jacob (Eds.), Aquatic functional biodiversity: An eco-evolutionary approach (pp. 75–111). Londres/San Diego, CA/Waltham, MA/Oxford: Academic Press (Elsevier).
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199, 1302–1310. doi: 10.1126/science.199.4335.1302
Connor, E. F., & McCoy, E. D. (2000). Species-area relationships. En S. A. Levin (Ed.), Encyclopedia of biodiversity (pp. 397–411). Nova York, NY: Elsevier. doi: 10.1016/B0-12-226865-2/00252-2
Hillebrand, H. (2004). On the generality of the latitudinal diversity gradient. The American Naturalist, 163(2), 192–211. doi: 10.1086/381004
Hortal, J., De Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., & Ladle, R. J. (2015). Seven shortfalls that beset large-scale knowledge of biodiversity. Annual Review of Ecology, Evolution, and Systematics, 46, 523–549. doi: 10.1146/annurev-ecolsys-112414-054400
Mittelbach, G. G. (2012). Community ecology. Sunderland, MA: Sinauer Associates.
Mittelbach, G. G., Steiner, C. F., Scheiner, S. M., Gross, K. L., Reynolds, H. L., Waide, R. B., … Gough, L. (2001). What is the observed relationship between species richness and productivity? Ecology, 82(9), 2381–2239. doi: 10.2307/2679922
Rosenzweig, M. L. (1997). Species diversity in space and time. Cambridge: Cambridge University Press.
Siqueira, T., Bini, L. M., Thomaz, S. M., & Fontaneto, D. (2015). Biodiversity analyses: Are aquatic ecologists doing any better and differently than terrestrial ecologists? Hydrobiologia, 750, 5–12. doi: 10.1007/s10750-014-2071-6
Sousa, W. P. (1979). Disturbance in marine intertidal boulder fields: The non-equilibrium maintenance of species diversity. Ecology, 60, 1225–1239. doi: 10.2307/1936969
Stein, A., Gerstner, K., & Kreft, H. (2014). Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology Letters, 17(7), 866–880. doi: 10.1111/ele.12277
Tilman, D., & Pacala, S. (1993). The maintenance of species richness in plant communities. En R. E. Ricklefs & D. Schluter (Eds.), Species diversity in ecological communities (pp. 13–25). Chicago: The University of Chicago Press.
Este trabajo ha recibido el apoyo de CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), del Ministerio de Educación de Brasil.




