

Evolución social
Una historia biológica de la cooperación

Hablar de la vida es hablar de cooperación. Su origen evolutivo, sus distintos niveles de organización o su actual complejidad son consecuencia de la cooperación entre entidades biológicas distintas. También lo son las sociedades animales, entre las que se incluye la sociedad más compleja de todas, la humana. Nuestro lenguaje, nuestra extraordinaria cultura, nuestras ciudades y vastas redes sociales, son fruto de la cooperación. ¿Cómo es posible que, en un mundo dominado por la competencia por los recursos, por la evolución darwiniana, la cooperación haya llegado a jugar un papel tan relevante? La evolución social, el estudio de las bases biológicas de la cooperación, nos permite responder a esta pregunta. En un recorrido que empieza con la primera célula y termina con la explosión de la vida social en los animales, la evolución social nos explica cómo y por qué la cooperación ha guiado la historia de la vida en este planeta.
Palabras clave: biología, cooperación, altruismo, comportamiento social, sociedades humanas, evolución social.
De átomos a dioses
Mucho antes de que el primer organismo vivo deambulara por este planeta aparecieron innumerables galaxias sembradas de inmensos motores termonucleares, las estrellas. En perpetua lucha entre la fuerza implosiva de su propia gravedad y la fuerza explosiva de las continuas fusiones nucleares de su interior, las estrellas sembraron el universo de los elementos de la tabla periódica. Entre ellos el carbono, base de los complejos compuestos orgánicos que encontramos en gases interestelares, asteroides y en las fuentes hidrotermales de nuestro fondo marino. En algún momento hace unos 3.800 millones de años, flotando en un mar de moléculas orgánicas al calor de estas fuentes hidrotermales, dichos «bloques de la vida» se organizaron para dar lugar a los primeros replicadores.
«El origen evolutivo de la célula está fundado en la cooperación»
En ese mismo instante, la evolución darwiniana comenzó a operar. De su mano, esa vida primigenia aprendió a organizarse en células y a saciar su incesante avidez de energía. Primero, con moléculas de su entorno y, después, con la energía inagotable del motor termonuclear más cercano, el sol. Una vez equipada con esos pequeños generadores de energía, la vida conquistó el planeta. Ocupó desde las fosas abisales hasta los picos más altos, desde el desierto más tórrido a la llanura más gélida. Aprendió a agruparse para formar, sucesivamente, organismos simbióticos, multicelulares y sociales. Aprendió, así, a cazar y a huir de otros organismos, a observar, oír, palpar y percibir las perturbaciones mecánicas, los fotones, los campos electromagnéticos y las sustancias químicas que hicieron tangible su mundo. Aprendió a correr, reptar, planear, volar, nadar y excavar, incluso a navegar usando las estrellas del firmamento. Aprendió a cantar, a acariciarse, a emocionarse, a amar. Aprendió a aprender, produciendo sistemas nerviosos lo suficientemente complejos como para recordar el pasado y anticipar el futuro. Aprendió a ser consciente, a usar y construir herramientas y, en un linaje insignificante de primates africanos, a usar símbolos para propagar su memoria de generación en generación. Por primera vez en este planeta, acaso en el universo, la vida se preguntó por sí misma, por el secreto de su propia existencia. Pintó la Mona Lisa, compuso la novena sinfonía y construyó enormes ciudades de calles reticulares y rascacielos asombrosos. Aprendió, incluso, a partir el átomo, a explorar el cosmos y a rediseñarse a sí misma: las hazañas de un dios.
Hay un hilo conductor a todos estos hitos, algo intrínseco a la vida; algo que no solo la ha acompañado en su insólito camino, sino que en gran medida la ha guiado (Bourke, 2011; Marshall, 2015).
El teatro evolutivo de la cooperación
Como sabemos, la vida de este planeta (y posiblemente la de muchos otros) se basa en la célula. Lo que resulta menos conocido es que el origen evolutivo de la célula está fundado en la cooperación. Situémonos aproximadamente entre 500 y 1.000 millones de años tras la formación de la Tierra (hace unos 3.500-4.000 millones de años). En ese periodo acontecen una serie de cambios críticos que hacen que los primeros replicadores, esas primeras moléculas con capacidad para reproducirse que ya podemos llamar vida, se organicen en una célula. Este hito es absolutamente clave, pues conforma el primer «individuo» y, con ello, la aparición de los procariotas (i. e., bacterias y arqueas). De todos los fenómenos que tuvieron que darse, uno de los más sorprendentes fue que distintos replicadores (los futuros genes) cooperasen para organizarse en un mismo genoma envuelto por una membrana. Es más, posiblemente, la aparición del primer individuo vivo propiamente dicho aceleró la acción de la selección natural, un proceso evolutivo ya irreversible que terminaría por transformar la superficie del planeta, por aquel entonces aún yermo e inhóspito.

Una de las consecuencias últimas de la multicelularidad es la aparición de las primeras sociedades animales. Multitud de organismos se organizan en grupos sociales, como es el caso de los lobos o las colonias coralinas./ Eva Blue – Unsplash
Como consecuencia de este proceso, tras la aparición de los primeros procariotas, la vida sufrió una explosión en su diversidad (una radiación adaptativa) de la que surgieron multitud de especies diferentes de estos microorganismos, cada una adaptada a explotar un aspecto distinto del ambiente primigenio en el que vivían. Tras cientos de millones de años de evolución en este mundo procariota, ocurrió uno de los fenómenos más determinantes en la historia de la vida: un procariota (una bacteria) fue «engullido» por otro (una arqueobacteria) y ambos se fusionaron en un nuevo organismo por medio de un proceso denominado «endosimbiosis». Esta fusión constituyó todo un acontecimiento: dos células sencillas cooperando estrechamente para dar lugar a un nuevo tipo de individuo, con un núcleo definido1 y una ventaja extraordinaria. Esa pequeña célula «engullida» se especializó en la mitocondria, un orgánulo celular abundante en las actuales células eucariotas. Las mitocondrias actúan como centrales energéticas, proporcionando mucha más energía de la que es capaz de generar una sola célula procariota (algunas células cuentan con millares de mitocondrias), lo cual permitió a la célula eucariota crecer en tamaño y complejidad. Más energía disponible supone un metabolismo mayor y, por lo tanto, la capacidad para realizar más trabajo. Entre otras cosas, esto permitió incrementar el tamaño del genoma y sintetizar más proteínas, combinándolas en multitud de formas para crecer en complejidad y colonizar nichos ecológicos inaccesibles a los procariotas. Sabemos, además, que la endosimbiosis se repitió al menos otra vez (probablemente alguna más) para dar lugar a los cloroplastos, las «centrales solares» que hacen posible la fotosíntesis y que dieron lugar a las plantas y las algas que tiñeron de verde la Tierra.
Así, procariotas y eucariotas empezaron a convivir en un mundo darwiniano donde la competencia por contribuir con más genes a las siguientes generaciones hace que la reproducción sea uno de los procesos bajo mayor presión de selección. Esos primeros organismos, como todos los procariotas actuales, se reproducían de forma asexual; generaban copias idénticas de su genoma que trasladaban, junto con parte de su citoplasma, a sus descendientes. Sin embargo, a pesar de no haberse hallado una huella clara en los fósiles, diversos análisis filogenéticos sugieren que en algún ancestro común a todos los eucariotas se originó una forma enigmática y más divertida de reproducirse, el sexo. Una mayoría abrumadora de organismos eucariotas, se estima que más del 99,9%, se reproducen sexualmente. El sexo implica que dos organismos distintos generen gametos en los que solo están representados la mitad de sus genes y se fusionen para formar un zigoto. Es decir, el sexo requiere que dos organismos renuncien a la mitad de sus genes en pro de la formación de un descendiente común. De nuevo, cooperación.
La siguiente gran transición evolutiva daría lugar a un nuevo tipo de individuo y también estaría basada en la cooperación: la multicelularidad. Sabemos que la transición de organismos unicelulares a organismos multicelulares ha sucedido de forma independiente al menos 25 veces en el árbol de la vida. En cada una de estas ocasiones, diversas células debieron organizarse de forma cohesiva para dar lugar a un solo individuo. Esta cooperación ha de ser tan estrecha que la mayor parte de las células que componen un organismo multicelular sencillamente renuncian a perpetuar sus genes; solo un reducido número de células, las germinales, se reproducen. Las ventajas de la multicelularidad son indudables. Incluyen, por ejemplo, la capacidad de crecer mucho más allá de los límites físicos que imponen un tamaño celular máximo, y abre la posibilidad a la diferenciación y especialización celular. En definitiva, da lugar a individuos mucho más complejos que pueden acceder a nichos ecológicos hasta entonces inalcanzables para la vida. De hecho, con la multicelularidad se suceden diversas radiaciones evolutivas que terminan por dar lugar a los grandes grupos de eucariotas multicelulares; los hongos, animales, algas y plantas que, junto a los procariotas, conforman la totalidad de nuestro bioma.
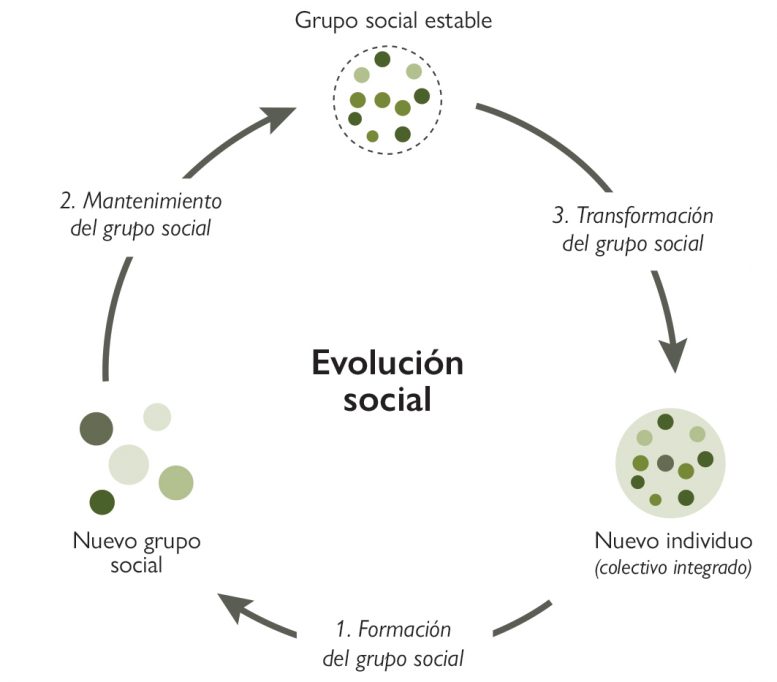
Figura 1. Fases en la evolución social de un nuevo individuo. 1) Individuos distintos cooperan entre sí para obtener beneficios directos y/o indirectos, dando lugar a un nuevo grupo social; 2) evolucionan mecanismos sociales que estabilizan el grupo, favoreciendo la cooperación; 3) por último, los mecanismos de cohesión son tan eficaces que se produce una transición hacia un nuevo colectivo integrado, un nuevo individuo o nivel de organización biológica.
Una de las consecuencias últimas de la multicelularidad es la aparición de las primeras sociedades animales. En las colonias de invertebrados marinos como los corales, en la bucólica estampa de una manada de lobos aullando a la luz de la luna, en el agitado termitero, multitud de organismos se organizan en grupos sociales. La vida social de los animales exhibe una enorme variación en su complejidad y, por tanto, una enorme variedad en el grado de cooperación que implica. A veces, esta es pasajera o circunstancial, mientras que en otras ocasiones es tan estrecha que la supervivencia de todo el grupo depende de ella. Es el caso de muchos insectos eusociales, como las abejas o las susodichas termitas cuya división y especialización del trabajo conlleva que solo se reproduzca un subconjunto del grupo social (e. g., la abeja reina). Esta segregación de la labor reproductiva recuerda sin duda a la especialización de las células germinales en los organismos multicelulares. De hecho, tal es el grado de cohesión de algunos de estos grupos sociales que ciertos especialistas han llegado a presentarlos como una suerte de «superorganismos», como un nuevo tipo de individuo en la escala de organización biológica. En todo caso, la vida social se apoya en la cooperación. En algunas especies, la sociabilidad ha permitido trasladar conocimiento crítico de generación en generación por medio del aprendizaje, prescindiendo de los genes. Este fenómeno, que denominamos cultura, no es un coto privado de la especie humana. Animales tan diversos como las hormigas, los carboneros, los macacos japoneses, el pez espinocho o las orcas transmiten información cultural relativa a las técnicas de forrajeo, las predilecciones a la hora de escoger pareja, las formas de evadirse de los depredadores, las preferencias alimentarias o las rutas de migración por medio de un aprendizaje social transmitido a lo largo del tiempo (Laland, 2008). Obviamente, la cultura ha revelado su verdadero poder en la especie humana, ayudándonos incluso a escapar en cierta medida de la selección natural. En última instancia, la cultura nos ha enseñado a establecer redes de cooperación a escala global, permitiéndonos esculpir el mundo, e incluso la vida, a nuestro antojo.
¿Cómo es posible? ¿Por qué, en un mundo dominado por la implacable competencia darwiniana, resulta tan ubicua la cooperación? ¿Qué explica que incluso los distintos niveles de organización de la vida reflejen grandes transiciones evolutivas basadas en la cooperación (Figura 1)?
Altruismo: «Mi vida por dos hermanos»
Nos encontramos a finales de los años cuarenta o principios de los años cincuenta. En el ya derruido pub The Orange Tree, próximo al University College de Londres, un grupo de académicos departe amigablemente sobre ciencia. Súbitamente, uno de los tertulianos, corpulento, de poblado bigote y cejas eternamente fruncidas, abandona la conversación y, durante unos instantes, garabatea cálculos en una servilleta. Se trata de J. B. S. Haldane, Jack para amigos y familiares, uno de los padres de la genética de poblaciones y una figura clave en la biología evolutiva del siglo XX2. Al acabar, declara con su voz sonora e inconfundible: «Daría con gusto mi vida por dos hermanos u ocho primos».
Esa sencilla frase, motivo de esta curiosa anécdota narrada por el también célebre John Maynard Smith, por entonces pupilo de Haldane, esconde una de las ideas más acertadas, influyentes y poderosas de la biología evolutiva reciente. No era una novedad que, en cualquier especie, unos individuos están más estrechamente emparentados que otros y, por lo tanto, comparten una mayor cantidad de genes. Lo que Haldane intuyó es que este hecho tan incontrovertido puede explicar la evolución de un tipo de comportamiento difícil (hasta entonces) de casar con la selección natural, el altruismo. El sacrificio de una araña social africana (Stegodyphus dumicola), que se deja devorar por su descendencia; las llamadas de alarma que profieren los perrillos de las praderas (Urocitellus beldingi) para alertar del peligro de un depredador a sus congéneres, poniendo en riesgo su propia supervivencia, o el trabajo incesante de una abeja obrera para criar las larvas de su abeja reina, son casos de altruismo. En todos ellos, hay individuos con comportamientos que aparentemente reducen su propia eficacia biológica (i. e., su supervivencia o su reproducción) para favorecer la de otros de su especie. Por definición, el altruismo alude a un tipo de cooperación que no parece reportar beneficio directo alguno ni a corto ni a largo plazo, y por consiguiente parece desafiar la lógica evolutiva.
Nada más lejos de la realidad. Aunque Haldane no profundizaría en esta idea, la lógica evolutiva del altruismo quedaría formalizada matemáticamente por el brillante W. D. Hamilton en 1964 (Hamilton 1964a y 1964b), y fue bautizada por John Maynard Smith como la «selección por parentesco» (Maynard Smith, 1964). La idea, en breve, queda encapsulada por la conocida «regla de Hamilton»:
rB > c
Un comportamiento con coste c para el actor será igualmente favorecido evolutivamente si los beneficios para el individuo receptor B, multiplicados por el grado de parentesco entre actor y receptor r, son mayores. En otras palabras, los comportamientos altruistas tienen sentido evolutivo porque los individuos pueden favorecer la representación de sus genes en las generaciones siguientes por dos vías: reproduciéndose ellos mismos (i. e., de forma directa) o favoreciendo la reproducción de aquellos que comparten sus genes (i. e., de forma indirecta). Lo que Haldane quería decir es que la reproducción de dos de sus hermanos, con los que compartiría en promedio un 50% de sus genes en cada caso, era equivalente en términos evolutivos a su propia reproducción. Haldane transferiría la mitad de sus genes a un hijo/a, la misma cantidad de sus propios genes que, en promedio, comparte con dos de sus sobrinos. Así pues, Hamilton entendió que, en realidad, la evolución favorece aquellas adaptaciones que tienden a maximizar la suma de la transmisión directa e indirecta de genes a las futuras generaciones, lo que denominó «eficacia inclusiva». En muchas ocasiones, esto implica que la mejor estrategia que puede seguir un organismo para transmitir más copias de sus genes a las siguientes generaciones consiste en sacrificar su propia reproducción en beneficio de la de sus parientes. Este principio, tan aparentemente sencillo, nos ha permitido explicar desde la evolución del altruismo en los animales sociales hasta algunas de las grandes transiciones evolutivas que, como la multicelularidad, denominamos transiciones «fraternales» aludiendo precisamente a la selección por parentesco (Bourke, 2011). En las transiciones fraternales, distintos individuos (e. g., células individuales) cooperan entre sí para cohesionarse y formar un nuevo individuo (e. g., un organismo multicelular) por los beneficios indirectos (i. e., en términos de eficacia inclusiva) que esta colaboración conlleva. El posible conflicto evolutivo que afrontan las distintas células de un organismo multicelular, donde todas trabajan en concierto para que solo unas pocas se reproduzcan, es mínimo porque todas las células están estrechamente emparentadas unas con otras; en nuestro caso, como en el de la mayor parte de organismos multicelulares, todas las células de nuestro cuerpo son (salvo mutaciones) genéticamente idénticas.
Beneficios directos
Los murciélagos vampiro (Desmodus rotundus) son animales fascinantes. Sus hábitos alimenticios han inspirado innumerables mitos e historias, tanto que han llegado a formar parte de un imaginario colectivo casi global. En sus salidas nocturnas, estos inofensivos mamíferos hematófagos buscan ganado o ungulados salvajes. Cuando los localizan, usan sus incisivos para realizar una pequeña herida de la que extraen una cantidad de sangre (unos 25 ml a la hora) insignificante para sus «víctimas». De hecho, esta inusual fuente de alimento es tan escasa y poco nutritiva que morirán de inanición si no consumen sangre aproximadamente cada tres días. Dada su extraordinaria longevidad (hasta 18 años), esto significa que los vampiros tienen que asegurarse una comida caliente casi a diario, toda una proeza si tenemos en cuenta que sus fuentes de alimento son poco previsibles. La solución a este problema común radica en la cooperación.

Durante el día, los vampiros se refugian en cuevas o troncos huecos de los árboles, en grupos sociales muy estables. Cuando un vampiro regresa al grupo después de una noche infructuosa, otros miembros de su grupo regurgitan una cantidad de sangre suficiente para aplacar su apetito. En contraprestación, este devolverá el favor en ocasiones futuras. De hecho, las donaciones no son aleatorias, sino que tienden a producirse entre vampiros con vínculos sociales estrechos, y el factor que mejor predice la cantidad de comida que se va a compartir con otro murciélago es la cantidad de comida recibida de él en el pasado. Este es uno de los casos clásicos y mejor documentados de un fenómeno que se denomina «altruismo recíproco», postulado en 1971 por el tan brillante como extravagante etólogo Robert Trivers.3 Para que el altruismo recíproco sea estable evolutivamente tienen que cumplirse tres premisas. En primer lugar, tiene que darse la oportunidad de que los mismos individuos interaccionen repetidamente a lo largo del tiempo, para que ambos puedan salir beneficiados de la cooperación. En segundo lugar, los beneficios de recibir ayuda deben ser mayores que los costes de proporcionarla. En tercer lugar, los individuos deben ser capaces de reconocer individualmente a sus congéneres para evitar compartir alimento con aquellos individuos que no cooperen (impidiendo que se propague la estrategia egoísta). Todas estas condiciones se cumplen en el caso de los vampiros y, qué duda cabe, también subyacen en muchos de los comportamientos cooperativos de nuestra especie.
En realidad, el altruismo recíproco no es sino uno de los distintos tipos de mecanismos mediados por unos beneficios directos que sirven para mantener la cooperación entre los miembros de un grupo social. Lo que tienen en común todos los tipos de cooperación de esta clase es que el donante (i. e., el que ayuda) obtiene de ella unos beneficios que, a corto o medio plazo, redundan en el incremento de su propia eficacia directa. El problema es también el mismo en todos los casos, y consiste en cómo impedir que se propague la estrategia egoísta de no cooperar y sacar provecho de la cooperación de otros. Aquí entran en juego diversos mecanismos sociales de control que, como en la especie humana, tienden a premiar la cooperación y a castigar el egoísmo (Riehl y Frederickson, 2015). Por ejemplo, en algunas especies de aves, peces e insectos con cría cooperativa, los ayudantes subordinados son acosados o castigados por los dominantes cuando no contribuyen. No obstante, parece que en la mayor parte de los casos, los beneficios que a corto o largo plazo reporta la ayuda constituyen un incentivo evolutivo suficiente como para mantener la cooperación. En el caso de la cría cooperativa, los contribuyentes no reproductores suelen incrementar su supervivencia y sus habilidades para criar con éxito en el futuro.
«La evolución social estudia cómo y por qué la cooperación ha dado lugar a todo tipo de comportamientos y adaptaciones sociales»
Este mismo principio, el de los beneficios directos mutuos, explica la cooperación entre especies diferentes. Cuando un pez limpiador escoge a su cliente, este, un depredador mucho mayor que podría engullirlo con facilidad, le dejará acercarse y alimentarse de los parásitos, la piel muerta y el tejido infectado de sus branquias y su superficie corporal. Es esta una relación mutualista de la que ambos extraen beneficios críticos para su supervivencia y reproducción. El primero depende de su cliente como fuente de alimento y el segundo necesita al otro para librarse de parásitos y mantener su piel sana. Las relaciones de este tipo con frecuencia llegan a extremos tales que ambas especies terminan por ser completamente dependientes la una de la otra. En ocasiones, incluso dan lugar a la formación de un nuevo tipo de individuo. Tal es el caso de los líquenes, fruto de la cooperación estrecha entre un hongo y una o varias poblaciones de algas o cianobacterias. Es este, también, el telón de fondo de los procesos de endosimbiosis que dieron lugar a la célula eucariota y que, como hemos visto, constituyen uno de los pasos más determinantes de la historia de la vida.
Conclusiones
El 27 de diciembre de 1831 el H. M. S. Beagle zarpó del puerto de Plymouth rumbo a Tierra del Fuego en un viaje que, a lo largo de cinco años, llevaría a un entonces jovencísimo Charles Darwin, recién graduado por la Universidad de Cambridge a sus 22 años, alrededor del globo. Desde Suramérica a las islas del Pacífico pasando por Tahití, Australia, Nueva Zelanda, África y muchas de las islas del Atlántico, esta aventura cambiaría la vida del naturalista inglés y marcaría la historia de la ciencia y de la humanidad. En su delicioso diario de abordo, Darwin relata los pormenores de una expedición que le llevaría a recopilar centenares de especímenes de animales y plantas, como los famosos pinzones de las islas Galápagos. Escala a escala, anotación a anotación y espécimen a espécimen, el germen de una idea fue creciendo en su mente, una idea que tardaría años en elaborar, madurar y perfeccionar. Nada más y nada menos que 28 años transcurrirían desde que el Beagle levara anclas en aquel frío día de diciembre hasta la publicación de su opus magnum, El origen de las especies, el 24 de noviembre de 1859 (Darwin, 1859). La primorosa labor de Darwin, recopilando y describiendo la extraordinaria diversidad de formas, colores y comportamientos que se encontró durante este viaje, y durante muchos años después en sus posteriores investigaciones, terminaría por alumbrar una de las teorías más brillantes, revolucionarias y (para muchos) peligrosas de la historia. En palabras del filósofo Daniel Dennet (1995): «[la peligrosa idea de Darwin] corroe casi todo concepto tradicional […] es un solvente universal, capaz de golpear al corazón de todo lo que alcanza la vista. La cuestión es, ¿qué deja atrás?».
Más de un siglo después de la muerte de Darwin, y sobre todo a partir de mediados del siglo XX, comenzamos a entender que una de las cuestiones sobre las que opera ese «solvente universal» es la cooperación, dando lugar a una de las disciplinas científicas sin duda más excitantes y ambiciosas de la biología evolutiva: la evolución social. Su objeto de estudio es cómo y por qué la cooperación ha dado lugar a todo tipo de comportamientos y adaptaciones sociales, incluidas algunas (si no todas) de las grandes transiciones evolutivas que han moldeado la vida. Es la disciplina a la que acudimos para intentar entender desde las relaciones más sencillas, como la cooperación entre bacterias, hasta los aspectos biológicos de la cooperación humana, pasando por la formación de nuevos tipos de individuos y escalas de complejidad biológica. En definitiva, es el estudio científico de la evolución y función de la cooperación. Algo tan fundamental que, sin ello, resulta imposible entender la vida, desde su origen en esos primeros replicadores al calor de las fuentes hidrotermales hasta la maraña de relaciones sociales que conforman nuestra sociedad y nuestra cultura.
Notas
1. De ahí su nombre, eucariotas (eu, “verdadero”; karyon, “núcleo”). (Tornar al text)
2. Descrito famosamente como «un enorme rinoceronte lanudo de temperamento imprevisible», y a pesar de no disponer de título oficial alguno en ciencias, Haldane era un erudito en física, química y, por supuesto, en biología, además de un gran amante de la poesía. Sus colegas dijeron de él que era el hombre más inteligente de su generación, el biólogo más erudito del siglo XX, e incluso que «parece el último hombre que posiblemente sabe todo lo que hay que saber». También decían que nunca renunciaba a una buena pelea y que jamás nadie bebió más que él y siguió sentado a la mesa para contarlo. (Tornar al text)
3. En sus impagables memorias (Trivers, 2017), este relata, entre muchas otras de sus aventuras, el tiempo que pasó entre rejas, su papel en la fundación de un grupo armado para defender a los homosexuales en Jamaica, o la vez que ayudó a huir a Huey Newton, el entonces líder de los panteras negras. (Tornar al text)
Referencias
Bourke, A. F. G. (2011). Principles of social evolution. Oxford Series in Ecology and Evolution. Oxford University Press.
Darwin, C. (1859). The origin of species. Murray & Sons.
Dennet, D. C. (1995). Darwin’s dangerous idea. Simon & Schuster.
Hamilton, W. D. (1964a). The genetical evolution of social behaviour. I. Journal of Theoretical Biology, 7, 1–16. https://doi.org/10.1016/0022-5193(64)90038-4
Hamilton, W. D. (1964b). The genetical evolution of social behaviour. II. Journal of Theoretical Biology, 7, 17–52. https://doi.org/10.1016/0022-5193(64)90039-6
Laland, K. (2008). Animal cultures. Current Biology, 18(9), R366–R370.
https://doi.org/10.1016/j.cub.2008.02.049
Marshall, J. A. R. (2015). Social evolution and inclusive fitness theory. Princeton.
Maynard Smith, J. (1964). Group selection and kin selection. Nature, 201, 1145–1147. https://doi.org/10.1038/2011145a0
Riehl, C., & Frederickson, E. (2015). Cheating and punishment in cooperative animal societies. Philosophical Transactions of The Royal Society B, 371, 20150090. https://doi.org/10.1098/rstb.2015.0090
Trivers, R. (2017). Vida indómita. Antoni Bosch.




