

Sobrevivir a la incertidumbre
Biodiversidad, adaptación y fluctuación ambiental en rotíferos

El estudio de la evolución frente a la incertidumbre ambiental es crucial para entender la diversidad biológica, ya que diversificar las estrategias de vida es clave para sobrevivir y reproducirse en un ambiente incierto. Los rotíferos son microinvertebrados planctónicos que habitan masas de agua continentales. Su ciclo vital complejo, que combina la reproducción sexual y la asexual, junto con un pequeño tamaño y tiempos de generación cortos, los convierte en excelentes organismos modelo en estudios de ecología evolutiva. Aquí presentamos estudios poblacionales de campo y de evolución experimental en el laboratorio donde mostramos cómo estos organismos son capaces de adaptarse localmente diversificando su ciclo vital frente a fluctuaciones impredecibles en su ambiente.
Palabras clave: plancton, rotíferos, ciclos vitales complejos, ambientes temporales, impredecibilidad ambiental.
«Si un organismo fuera capaz de adaptarse a cualquier circunstancia presente en la biosfera, sería el único ser vivo en la Tierra»
Introducción
Si un organismo fuera capaz de adaptarse a cualquier circunstancia presente en la biosfera –es decir, pudiera subsistir en los trópicos y en los polos, en el agua y en la atmósfera, a partir de cualquier fuente biótica o abiótica de energía– sería probablemente el único ser vivo en la Tierra y la diversidad de organismos en esta sería mínima. Contrariamente, en la práctica, adaptación significa especialización, e implica mejorar las aptitudes en relación con unas condiciones determinadas, sacrificando unas capacidades por otras. Por eso, una «dimensión», pero no la única, para explicar la biodiversidad es la adaptación. La adaptación se requiere para la persistencia de una especie en una localidad, y en consecuencia en un hábitat o en una región, y en la biosfera. El darwinismo nos enseña que la adaptación ocurre por selección natural. Esta, como cualquier otra fuerza natural, actúa como lo que un aristotélico llamaría «causa eficiente»,1 desde el pasado y sin finalidad. Es decir, cuando un ser vivo se encuentra ante un ambiente que de alguna manera es «genuinamente» nuevo, no cabe esperar que esté adaptado a él. En el mejor de los casos, entrará en un proceso de selección natural que acabe adaptándolo a la novedad. Dado que todos los ambientes son cambiantes, esto representa un reto crucial para el ajuste entre los organismos y su ambiente.
«Adaptación significa especialización, e implica mejorar las aptitudes en relación con unas condiciones determinadas»
Para darse cuenta de la ubicuidad de los cambios ambientales basta pensar en los propios del clima, y reparar en lo que guardamos en nuestros armarios (ropa de verano, de invierno, de entretiempo…) o considerar nuestras dudas sobre qué llevar cuando salimos de excursión. Estos dos ejemplos ilustran casos de «fluctuaciones ambientales», en los que existe una recurrencia frecuente en las condiciones, y que contrastan con las «novedades ambientales», como podría suponer un enfriamiento del clima en escala temporal geológica o la irrupción de un nuevo patógeno. Manteniendo el símil: no guardamos ropa en el armario en previsión de una época glaciar. En el caso de las fluctuaciones, la recurrencia se da dentro de la escala temporal propia de la respuesta evolutiva de los organismos, y puede actuar como una presión selectiva por la que evolucione la adaptación no solo a un valor promedio de las condiciones ambientales, sino a la variabilidad de estas.
«Si la adaptación a la incertidumbre ambiental es importante en cualquier circunstancia, será probablemente crucial en el futuro»
En este artículo tratamos la adaptación a la fluctuación ambiental, pero no de cualquier tipo. Los ejemplos del armario y la excursión ilustran dos modos muy diferentes de fluctuación. Algunas son, bien por su periodicidad, bien porque hay indicios que las anticipan, predecibles para los organismos. Otras no lo son. Aun así, la media y la varianza (esta última, la medida estadística más común para cuantificar la variación) de un factor que fluctúa impredeciblemente pueden bastar para que, por selección natural, aparezca una optimización en los rasgos del organismo, (optimización en el sentido de que maximiza la eficacia biológica, a veces denominada aptitud). La teoría evolutiva así lo apunta, y nuestros estudios empíricos, como veremos, también. Pero antes, advirtamos de una complicación de segundo orden: una de las novedades ambientales que cabe esperar como consecuencia del cambio global es el incremento de las fluctuaciones y su impredecibilidad. Si la adaptación a la incertidumbre ambiental es importante en cualquier circunstancia, será probablemente crucial en el futuro.
Estrategias de minimización de riesgos
Para un organismo que necesariamente cumple su ciclo vital en un año, de nada le sirve dejar muchos descendientes la mayor parte de los años si en otros no puede dejar ninguno. Esos annus horribilis representarían para él una barrera infranqueable. Su eficacia biológica, considerando el conjunto de años, sería cero. Por tanto, la media aritmética de la eficacia biológica en las diferentes situaciones ambientales, a menudo utilizada para describir el éxito reproductivo individual, no es en este caso una medida adecuada. En ambientes impredecibles, esperamos adaptaciones que hagan que los individuos no maximicen su eficacia promedio; las predicciones evolutivas no pueden hacerse sin tener en cuenta la varianza en la eficacia. Así, cuando hay una necesidad de anticiparse a cambios impredecibles, se espera la evolución de estrategias de minimización de riesgos (bet hedging) que reducen la varianza de la eficacia a costa de disminuir la media aritmética de la eficacia, pero que aumentan su media geométrica; es decir, maximizan la eficacia biológica a largo plazo.
«Por la escala temporal a la que opera, la respuesta adaptativa de los organismos no es fácil de estudiar»
La teoría evolutiva ha investigado este problema con diversas aproximaciones. Una simple, pero que a pesar de ello captura lo esencial del problema, supone que un organismo puede enfrentarse a dos tipos de condiciones (favorables o adversas), las cuales se suceden con cierta probabilidad en una serie impredecible. Un modo de minimizar los riesgos es poseer unas características constantes que funcionen bien cualesquiera que sean las condiciones. Estas características permitirían a la especie persistir en las condiciones adversas a costa de no explotar completamente las condiciones favorables siguiendo una estrategia conservadora. Otro modo de minimizar riesgos es producir cierta fracción de descendientes ajustada a las condiciones adversas y cierta fracción ajustada a las favorables. Se trata de aumentar las opciones de éxito con una estrategia diversificadora. Así, Cohen (1966) analizó el problema de una planta anual que pasa el invierno en forma de semilla, la cual germina al acercarse la primavera sin información sobre si la calidad del año permitirá alcanzar la fase de floración y fructificación. Su predicción fue que en estas plantas una fracción de las semillas producidas por adulto debería germinar (por si el año es bueno), mientras que otra fracción debería mantenerse en latencia (por si el año es malo), y este esquema de fraccionamiento debería repetirse año tras año para las semillas que han sobrevivido en latencia el año anterior. La existencia de bancos de semillas latentes en los suelos es un indicio que apoya esta predicción.
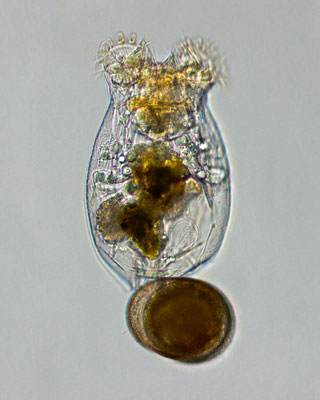
Figura 1. Microfotografía de una hembra de rotífero con un huevo de resistencia. Estos diminutos invertebrados suelen formar parte del plancton de lagos y lagunas, y su ciclo vital complejo los convierte en excelentes organismos modelo en estudios de ecología evolutiva. / Ivana Jezkova
Los rotíferos, organismos modelo en ecología y evolución
Por la escala temporal a la que opera, la respuesta adaptativa de los organismos no es fácil de estudiar. O se recurre a inferencias indirectas o se requieren sistemas experimentales especiales. Uno de estos sistemas lo proporcionan los rotíferos (Serra, García-Roger, Ortells y Carmona, 2019), unos pequeños invertebrados (0,1-0,4 mm de longitud; Figura 1) que con frecuencia forman parte del plancton de lagos y lagunas. Su característica más interesante es la partenogénesis cíclica, el ciclo vital de muchos de ellos. La partenogénesis cíclica (Cuadro 1) consiste en la combinación de la reproducción sexual y la asexual. Esta última, también llamada partenogénesis –literalmente, reproducción por vírgenes2–, se basa en la producción de hembras que, salvo mutación, son copias genéticas idénticas de sus madres. Sin necesidad de apareamiento e inversión en machos, la reproducción asexual, al combinarse con el tamaño corporal pequeño de estos organismos, da tasas de proliferación elevadas (la población puede duplicarse en un par de días), que permiten una colonización rápida del hábitat y tamaños poblacionales grandes. Esto es algo crítico debido a que el hábitat de los rotíferos es utilizable tan solo durante un periodo del ciclo anual, la llamada «estación de crecimiento», tras la cual la población activa desaparece. Antes de que llegue el final de la estación de crecimiento debe ocurrir al menos una generación sexual, lo que en muchas especies se induce por hacinamiento a una determinada densidad poblacional. Entonces, nacen machos que copulan con hembras, cuyos huevos entran en latencia y resisten en el sedimento de la laguna en condiciones en las que la supervivencia de individuos activos es imposible. Comienza entonces la estación de latencia, que puede ser prolongada. El momento de la reproducción sexual es, pues, crítico, suficientemente tardío como para no comprometer la proliferación asexual y suficientemente temprano como para poder formar los huevos resistentes a tiempo. Claro que no hay dos años iguales en cuanto a la duración de la estación de crecimiento. Esa duración es incierta.
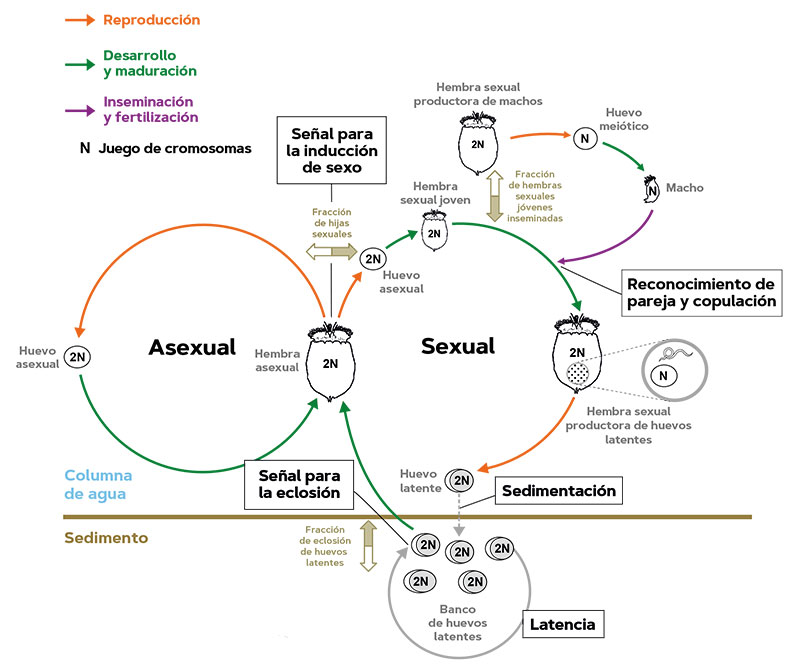
Cuadro 1. Las poblaciones de los rotíferos partenogenéticos cíclicos son típicamente temporales en el plancton y recolonizan la columna de agua durante la estación de crecimiento planctónico mediante la eclosión de huevos latentes de los sedimentos de las lagunas y los lagos. Los recién nacidos son hembras asexuales que producen huevos diploides (con dos juegos cromosómicos) que –salvo mutaciones– se desarrollan en hijas genéticamente idénticas a sus madres. Esta reproducción exclusivamente asexual se repite un número indeterminado de veces (proliferación clonal). La fase sexual –que no detiene completamente la reproducción asexual– comienza con la producción partenogenética de hijas sexuales por parte de madres asexuales como una fracción de su descendencia en respuesta a factores ambientales inductores. Las hembras sexuales producen huevos con un único juego de cromosomas (haploides) resultante de la meiosis (división celular en la que la dotación genética se reduce a la mitad) que se desarrollan partenogenéticamente en machos haploides o, si las hembras sexuales jóvenes se aparean, sus huevos haploides son fertilizados y se convierten en huevos latentes diploides. Estos huevos se depositan en el sedimento, pueden sobrevivir en condiciones adversas y permiten a las poblaciones recolonizar la columna de agua cuando se reanudan las condiciones adecuadas en el hábitat y también dispersarse a otros hábitats. Después de recibir los estímulos apropiados, una fracción de los huevos latentes eclosiona en hembras asexuales, y comienza una nueva estación de crecimiento. La fracción de los huevos en latencia que no eclosiona se acumula y forma bancos de huevos en el sedimento. / Fuente: Modificado a partir de Serra et al. (2019).
«El ciclo vital de muchos rotíferos es la partenogénesis cíclica, que consiste en la combinación de la reproducción sexual y la asexual»
La partenogénesis cíclica representa un delicado ajuste entre las ventajas y desventajas de la reproducción sexual y su rasgo concomitante en esos animales: la latencia. Este ciclo vital merece por sí mismo estudio y se ha utilizado para la investigación de hipótesis evolutivas sobre las ventajas de la reproducción sexual, hipótesis sobre la divergencia entre poblaciones locales, e hipótesis ecológicas sobre la coexistencia de especies (Serra et al., 2019). Además, el ciclo vital de los rotíferos ofrece la oportunidad para distinguir fácilmente entre factores genéticos y ambientales, ya que la reproducción asexual genera clones de individuos genéticamente idénticos. Por último, las altas tasas de proliferación asexual implican tiempos de generación cortos. Esto, junto al pequeño tamaño de los rotíferos, permite estudios de evolución experimental; es decir, estudios para comprobar si poblaciones de laboratorio responden a presiones de selección que imitan a las que se encuentran en su hábitat, pero que son creadas por el investigador en condiciones controladas.
Estudios con poblaciones ibéricas de rotíferos

Figura 2. En la imagen, muestreo en la laguna manchega de Hoya Rasa, incluidas en los estudios del Laboratorio de Ecología Evolutiva del Instituto Cavanilles de Biodiversidad y Biología Evolutiva de la Universitat de València. El muestreo de las poblaciones activas de rotíferos en la columna de agua se realiza con una red de zooplancton (en la imagen, al fondo) y el de sus formas latentes (huevos de diapausa) en el sedimento, con una draga Van Veen. En la misma imagen, al frente, un investigador recogiendo el sedimento extraído con la draga. / María José Carmona
El clima de la región mediterránea se caracteriza por variaciones irregulares que afectan al régimen de precipitaciones y evaporación. No es sorprendente que esta variación afecte a la hidrología de pequeñas masas de agua como las lagunas de la Mancha (Figura 2), en el este de la península ibérica. Para el estudio de esta región, Lluís Franch-Gras y colaboradores (Franch-Gras, García-Roger, Franch, Carmona y Serra, 2017), miembros del Laboratorio de Ecología Evolutiva del Instituto Cavanilles de Biodiversidad y Biología Evolutiva (ICBiBE) de la Universitat de València, adaptaron un conjunto de métodos de análisis de imágenes de satélite. El objetivo era la detección de masas de agua que son de área variable. Ello les permitió aprovechar una serie temporal de datos inusualmente larga en ecología. Mediante dicho análisis, se caracterizaron las irregularidades de los periodos anuales de inundación (el hidroperiodo) de las lagunas, lo que reveló la existencia de un rango amplio de impredecibilidad, desde lagunas muy predecibles hasta otras muy poco predecibles.
A partir de esas observaciones, pudo precisarse (Franch-Gras, García-Roger, Serra y Carmona, 2017) una primera hipótesis adaptativa, según la cual las poblaciones de rotíferos que viven en lagunas muy impredecibles tendrían un inicio de la reproducción sexual temprano, como una forma de producir huevos latentes también tempranamente y encarar con éxito una estación de crecimiento inesperadamente corta. Esto, no obstante, a costa de disminuir la reproducción asexual, y por tanto de explotar más plenamente una estación de crecimiento potencialmente larga. En una segunda hipótesis, muy cercana a la ya comentada para plantas anuales, estos investigadores propusieron que, en lagunas de hidroperiodo impredecible, los rotíferos poseerían genes que les permitirían producir una fracción alta de huevos latentes que no eclosionarían la primera vez que las condiciones del medio parecieran favorables. Así, se ahorrarían huevos que permanecerían latentes en el sedimento por si la estación de crecimiento fuese demasiado corta para completar el ciclo reproductivo con la producción de una nueva cohorte de huevos latentes, esto aun a costa de perder capacidades competitivas si la estación de crecimiento sí que permitiera completar el ciclo. Estas predicciones implican una diferenciación genética entre poblaciones de una misma especie, lo cual significa que se da una adaptación local. La comprobación de estas hipótesis debe hacerse preferentemente manteniendo los rotíferos procedentes de las distintas lagunas en condiciones de laboratorio, controlando los efectos ambientales no selectivos, pero que afectan al desarrollo de los organismos.
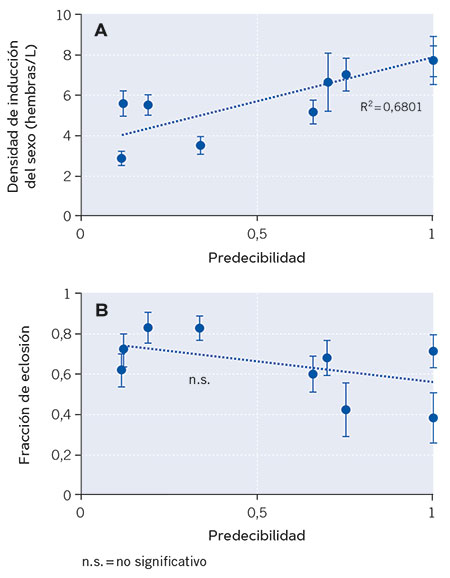
Figura 3. Relación entre dos rasgos relacionados con la latencia de nueve poblaciones naturales del rotífero partenogenético cíclico Brachionus plicatilis y el grado de predecibilidad ambiental de las lagunas que habitan. En la figura A, inicio de la reproducción sexual, inferido a partir de los valores de densidad poblacional a los que se induce. Un menor valor de densidad a la que se induce la reproducción sexual indica un inicio más temprano de este tipo de reproducción que garantiza la producción de huevos latentes en estos rotíferos. En la figura B, puede observarse la fracción de eclosión de huevos latentes (en tanto por uno). Una fracción menor garantiza la supervivencia si el ambiente es desfavorable. De cada rasgo se muestran los valores promedio para cada población y sus barras de error (± 1 error estándar). La línea discontinua en cada gráfica representa el ajuste por regresión mínimo-cuadrática. El coeficiente de determinación (R2 = 0,6801) es la proporción de la varianza en la densidad de inducción del sexo explicada por la predecibilidad ambiental. / Fuente: Modificado a partir de Franch-Gras et al. (2019).
Los resultados mostraron que, efectivamente, los genotipos procedentes de lagunas con impredecibilidad ambiental iniciaban la reproducción sexual más tempranamente que los procedentes de ambientes más predecibles (Figura 3a). Sin embargo, no se obtuvo evidencia a favor de la segunda hipótesis (Figura 3b), quizás porque basta con un rasgo –el momento de inicio de la reproducción sexual– para alcanzar la estrategia de minimización de riesgos necesaria, o quizás porque la heterogeneidad de condiciones en el sedimento es tan elevada que un rasgo intrínseco –como es la fracción de eclosión– es poco eficaz (Franch-Gras, García-Roger, Serra et al., 2017).
La evidencia proporcionada por los estudios realizados en nuestro laboratorio (Franch-Gras, García-Roger, Franch et al., 2017; Franch-Gras, García-Roger, Serra et al., 2017) es observacional, lo que implica que las relaciones se describen en su contexto natural –son realistas–, pero cabe preguntarse si esas relaciones podrían no ser causales; es decir, que los rasgos del ciclo vital se hubieran diferenciado entre poblaciones por alguna otra presión selectiva concomitante con la impredecibilidad ambiental. Para descartar esto se llevaron a cabo estudios de evolución experimental (Tarazona, García-Roger y Carmona, 2017). En el laboratorio, se crearon ambientes con fluctuación aleatoria en la longitud del hidroperiodo y ambientes con una longitud del hidroperiodo constante. En ellos, las poblaciones de rotíferos crecieron en una serie de ciclos de inundación. La biología de los rotíferos permitió simular esta complejidad en quimiostatos mantenidos en una cámara con condiciones controladas. Los resultados corroboraron las predicciones y además mostraron que la divergencia adaptativa esperada fue rápida. Las poblaciones de laboratorio evolucionaron hacia un inicio de la reproducción sexual más temprano en condiciones impredecibles, de acuerdo con la estrategia de minimización de riesgos conservativa apuntada por los resultados del estudio en poblaciones naturales (Figura 4a). Además, las poblaciones en el régimen impredecible también evolucionaron hacia fracciones de eclosión de huevos latentes menores que las del régimen predecible (Figura 4b). En otros estudios se ha reconocido la rapidez de evolución de los rotíferos (Declerck y Papakostas, 2016). La reproducción sexual combinada con una selección clonal que opera sobre todo el conjunto de los genes de un individuo son factores importantes. En nuestro sistema influye sin duda el que la población experimental inicial estaba compuesta de varios clones procedentes de diversas poblaciones naturales. A diferencia de las observaciones en el campo, en los estudios de evolución experimental se cumplieron las dos predicciones formuladas. Probablemente, las condiciones de laboratorio en las que eclosionan los huevos latentes, mucho más homogéneas que las de la naturaleza, hacen que el genotipo se exprese más limpiamente, y la fracción de eclosión sea un buen blanco para la evolución por selección.
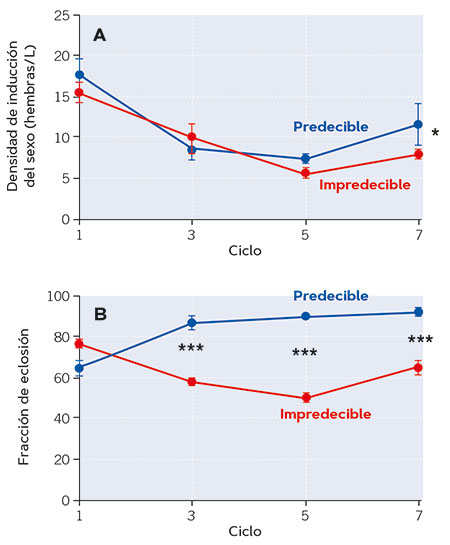
Figura 4. Evolución de dos rasgos relacionados con la latencia en poblaciones de laboratorio de Brachionus plicatilis a lo largo de siete ciclos de selección en regímenes divergentes de fluctuación ambiental en relación con la duración del hidroperiodo: predecible (duración constante del hidroperiodo a lo largo de los ciclos de crecimiento) e impredecible (hidroperiodo de duración variable a lo largo de los ciclos de crecimiento). En la figura A puede observarse el inicio de la reproducción sexual, inferida a partir de los valores de densidad poblacional a lo largo de los ciclos de evolución experimental. Un menor valor de densidad a la que se induce la reproducción sexual indica un inicio más temprano de esta. Así, por lo general, en condiciones más impredecibles, el inicio de la reproducción sexual suele ser más precoz. En la figura B, fracción de eclosión de huevos latentes (en porcentaje) a lo largo de los ciclos de evolución experimental. Puede observarse como, en los ciclos impredecibles, la fracción de huevos que eclosiona es menor (es decir, más cantidad se mantienen en latencia para mantenerse a salvo de condiciones potencialmente desfavorables). De cada rasgo se muestran los valores promedio para cada régimen y sus barras de error (± 1 error estándar). Los asteriscos indican diferencias estadísticamente significativas entre regímenes (*p < 0,05; **p < 0,01; ***p < 0,001). / Fuente: Modificado a partir de Tarazona et al. (2017).
El diseño de los estudios referidos (combinación de trabajo de campo y de laboratorio) permite conocer de forma no ambigua que las diferencias en los rasgos de ciclo vital son genéticas, es decir, que las diferencias no vienen confundidas por variaciones en el ambiente en que se expresan los genes. Por su parte, los resultados ofrecen una evidencia fuerte de que estas diferencias en el ciclo vital se deben a la selección natural en respuesta a la impredecibilidad ambiental; es decir, son debidas a una divergencia evolutiva adaptativa. Aun así, estos estudios no permitieron identificar los genes específicos responsables. En la actualidad, el desarrollo de las tecnologías de análisis genómico –en sus dos vertientes, la secuenciación del material genético y su análisis bioinformático– permite avanzar en ese último sentido incluso para especies que no han sido objeto de un estudio genético intensivo.
Así, en un nuevo estudio (Franch-Gras et al., 2018) se secuenció y elaboró un «borrador» de genoma para la especie estudiada, tras lo cual se analizó la divergencia entre los genomas de las poblaciones previamente caracterizadas por su adaptación diferencial a la impredecibilidad ambiental. Utilizando genotipado por secuenciación y el subsiguiente análisis bioinformático, se encontró un gran número de posiciones nucleotídicas en las que había divergencia (polimorfismos de nucleótido único, o SNP en sus siglas en inglés), incluso aplicando métodos muy conservativos. Este procedimiento también fue seguido por Tarazona y colaboradores para las poblaciones de laboratorio que habían divergido en los ensayos de evolución experimental. En ambos estudios, varios SNP –la mayoría de ellos ubicados dentro de genes– mostraron una diferenciación entre poblaciones mayor de lo esperado por azar, y se correlacionaron con la impredecibilidad ambiental y los rasgos del ciclo vital estudiados. Por ello, estos genes son candidatos a ser responsables de la adaptación local a ambientes impredecibles. Si bien la relación funcional de estos genes con los rasgos del ciclo vital no está esclarecida, representan un punto de partida para estudios que ya caen más en el campo de la biología molecular y celular que en el de la ecología evolutiva.
En su conjunto, los resultados muestran que las poblaciones de rotíferos pueden desarrollar respuestas adaptativas en rasgos relacionados con la latencia para ajustarse localmente al grado de impredecibilidad ambiental. Estas respuestas implican diferentes tipos de estrategias de minimización de riesgos. Los estudios llevados a cabo en el laboratorio de Ecología Evolutiva del ICBiBE proporcionan un análisis de la respuesta evolutiva de estas poblaciones, utilizando una aproximación integrada donde se cartografía el genotipo (marcadores genéticos) en el fenotipo (rasgos del ciclo vital), y el fenotipo en el ambiente (rasgos del hábitat).
«Existen fuerzas evolutivas que pueden oponerse a la selección natural y dificultar la adaptación»
Apareamiento preferencial y protección de la adaptación local
Existen fuerzas evolutivas que pueden oponerse a la selección natural y dificultar la adaptación. Para las poblaciones de rotíferos antes mencionadas, y que habitan lagunas muy cercanas entre sí, basta pensar qué efecto tendría sobre una población (la residente) la llegada de individuos procedentes de lagunas con diferente predecibilidad. Estos individuos no estarían adaptados a su nuevo destino, pero aun así podrían tener alguna oportunidad de, llegado el momento de la reproducción sexual, cruzarse con los residentes. El resultado de esa exogamia sería un difuminado de la divergencia adaptativa, y una pérdida de eficacia para la población residente. Para este escenario se ha postulado la llamada «hipótesis del refuerzo», muy importante en la teoría de la especiación (Butlin, 1987). En breve, el proceso descrito representa un cierto aislamiento reproductivo, ya que la descendencia exogámica no está adaptada a la localidad. Esto ocasionaría una presión de selección en contra de que residentes e inmigrantes llegaran a aparease, lo que reforzaría el aislamiento preexistente. Si el aislamiento se profundiza lo suficiente, las poblaciones divergentes llegarían a constituir especies diferentes, y así generarían diversidad biológica. En el Laboratorio de Ecología Evolutiva del Instituto Cavanilles, Ivana Jezkova desarrolla su tesis doctoral comprobando si los rotíferos de poblaciones locales divergentes muestran indicios de apareamiento preferencial intrapoblacional en razón de la impredecibilidad ambiental (Figura 5). Sus resultados preliminares apuntan en este sentido y, de confirmarse, mostrarían un mecanismo de estabilización de la divergencia adaptativa frente a la «perturbación» de la adaptación que significa la inmigración. Los resultados de este tipo son tremendamente interesantes porque la hipótesis del refuerzo no es trivial. En su contra está el hecho de que la reproducción sexual exige la coordinación de machos y hembras, su coincidencia temporal y espacial, y la identificación de pareja. Esa coordinación penaliza la variabilidad.

Figura 5. En el reconocimiento de pareja en los rotíferos intervienen quimiorreceptores, proteínas en la superficie corporal que actúan como señales mediante las cuales el macho (de tamaño menor que la hembra) reconoce a las hembras de su misma especie. En la imagen se observa el comportamiento de cortejo que incluye (de izquierda a derecha) el encuentro, giros alrededor de la hembra (circling) y la cópula. / Ivana Jezkova
«La reproducción sexual exige la coordinación de machos y hembras, su coincidencia temporal y espacial, y la identificación de pareja»
Conclusiones
La evolución por selección natural, si bien deja el registro de su acción en el material genético de los organismos, tiene uno de sus aspectos más llamativos en la adaptación. Esta solo puede entenderse completamente observando los rasgos morfológicos, fisiológicos y etológicos de los organismos pero, especialmente, observando también características del ciclo vital, dado que estas características están directamente relacionadas con la eficacia biológica (es decir, esas características son lo que se denomina «componentes de eficacia»). Libre de todo finalismo, esta teoría explica la adaptación por dos consistencias. Por un lado, la consistencia entre el ambiente pasado, que seleccionó determinados rasgos, y el ambiente presente, en el que se manifiesta el valor adaptativo de esos rasgos. Por otro lado, la consistencia en las características biológicas en los linajes, debida a la herencia. Esto convierte en más desafiante –y más fascinante de estudiar– la adaptación a la fluctuación ambiental, sobre todo si esta es irregular. La teoría, con su capacidad para idealizar procesos, apunta a que esa adaptación es en principio posible. Los experimentos de evolución llevados a cabo por el Laboratorio de Ecología Evolutiva muestran que se puede dar en seres vivos –como los rotíferos– comprometidos con muchas otras necesidades adaptativas. A su vez, los patrones descritos por los estudios de poblaciones naturales apuntan a que las causas identificadas en los experimentos actúan también en el entramado de factores que ocurre en la naturaleza. Quedan por esclarecer los mecanismos poblacionales de estabilización y –en otro nivel– los mecanismos moleculares y celulares subyacentes. Al respecto de esto último, en la era de las «-ómicas» (genómica, proteómica, transcriptómica, metabolómica…) se abre una tremenda oportunidad para sellar el hiato entre la biología molecular y la ecología evolutiva.
«Las poblaciones de rotíferos pueden desarrollar respuestas adaptativas para ajustarse localmente al grado de impredecibilidad ambiental»
Un mensaje final es que, frente a la incertidumbre ambiental, la evolución puede producir diversidad a distintos niveles: intraespecífica, que favorece la diversificación de las estrategias de vida (diversidad funcional) e interespecífica, que favorece los procesos de especiación (es decir, la diversidad clásica de especies).
1. En su Metafísica, Aristóteles describe cuatro clases de respuestas frente al porqué de las cosas. La causa material tiene que ver con el material del que están hechas las cosas (es decir, la causa de una mesa es la madera); la formal tendría que ver con su diseño (es decir, la forma, disposición o apariencia; por ejemplo, formalmente una mesa es una estructura plana apoyada sobre el suelo por patas); la causa eficiente tendría que ver con la interacción con otros agentes externos (en el caso de la mesa, la causa eficiente es el carpintero), y la causa final tendría que ver con la finalidad de las cosas, lo que invoca la existencia de un diseñador (es decir, la causa formal es la intención del carpintero de hacer una mesa para comer o estudiar en ella). (Vuelve al texto)
2. Del griego parthenos (“virgen”) y genesis (“creación”). (Vuelve al texto)
Parte del trabajo que se presenta se financió con fondos públicos del Ministerio Español de Economía y Competitividad (proyectos CGL2012-30779 y CGL2015-65422-P, ambos cofinanciados con fondos FEDER).
Referencias
Butlin, R. (1987). Speciation by reinforcement. Trends in Ecology & Evolution, 2(1), 8–13. doi: 10.1016/0169-5347(87)90193-5
Cohen, D. (1966). Optimizing reproduction in a randomly varying environment. Journal of Theoretical Biology, 12(1), 119–129. doi: 10.1016/0022-5193(66)90188-3
Declerck, S., & Papakostas, S. (2016). Monogonont rotifers as model systems for the study of microevolutionary adaptation and its eco-evolutionary implications. Hydrobiologia, 796(1), 131–144. doi: 10.1007/s10750-016-2782-y
Franch-Gras, L., García-Roger, E. M., Franch, B., Carmona, M. J., & Serra, M. (2017). Quantifying unpredictability: A multiple model approach for Mediterranean ponds by using satellite imagery. PLOS ONE, 12(11), e0187958. doi: 10.1371/journal.pone.0187958
Franch-Gras, L., García-Roger, E. M., Serra, M., & Carmona, M. J. (2017). Adaptation in response to environmental unpredictability. Proceedings of the Royal Society B: Biological Sciences, 284(1868), 20170427. doi: 10.1098/rspb.2017.0427
Franch-Gras, L., Hahn, C., García-Roger, E. M., Carmona, M. J., Serra, M., & Gómez, A. (2018). Genomic signatures of local adaptation to the degree of environmental predictability in rotifers. Scientific Reports, 8(1), 16051. doi: 10.1038/s41598-018-34188-y
Franch-Gras, L., Tarazona, E., García-Roger, E. M., Carmona, M. J., Gómez. A., & Serra, M. (2019). Rotifer adaptation to the unpredictability of the growing season. Hydrobiologia. doi: 10.1007/s10750-019-3886-y
Serra, M., García-Roger, E. M., Ortells, R., & Carmona, M. J. (2019). Cyclically parthenogenetic rotifers and the theories of population and evolutionary ecology. Limnetica, 38(1), 67–93. doi: 10.23818/limn.38.13
Tarazona, E., García-Roger, E. M., & Carmona M. J. (2017). Experimental evolution of bet hedging in rotifer diapause traits as a response to environmental unpredictability. Oikos, 126, 1162–1172. doi: 10.1111/oik.04186
Los cuatro autores forman parte del Laboratorio de Ecología Evolutiva del ICBiBE cuya investigación se centra en estudios de ecología evolutiva y ecología de poblaciones de microorganismos acuáticos, en concreto, en el análisis demográfico, genético y ecológico del zooplancton. Entre sus intereses científicos destacan la adaptación de los ciclos vitales de los rotíferos a ambientes variables, los procesos mediadores de la coexistencia de especies competidoras, la diferenciación poblacional y especiación, la latencia como estrategia de dispersión en el espacio y en el tiempo y los procesos evolutivos que mantienen la reproducción sexual en las poblaciones.




