

‘Homo sapiens’: ¿quiénes somos?
Características esenciales de nuestra especie

En este artículo analizamos cuáles son los rasgos que conforman a los humanos actuales a partir de su génesis. El objetivo es caracterizar la evolución biológica y cultural de la humanidad dentro del marco evolutivo de las especies de nuestro género. Jerarquizamos que características son las que nos distinguen dentro del reino animal: fabricar una gran diversidad de herramientas, el control del fuego, el lenguaje, el ritual funerario, etc. Tenemos que descubrir si realmente el aumento de complejidad solo se da en nuestra especie o se trata de un proceso por el que otras especies también han pasado o pasarán.
Palabras clave: hominización, humanización, singularidad, herramientas, lenguaje, complejidad, especie.
La singularidad de Homo sapiens puede representar la síntesis evolutiva de todo nuestro género y, por tanto, una expresión evolutiva singular. Somos una especie más, con un desarrollo y una complejidad que se han hecho patentes con la socialización de adquisiciones desarrolladas inicialmente por otras especies con las que hemos convivido o que nos han precedido. Consecuencia de esta captura de adquisiciones es el crecimiento exponencial que ha registrado nuestra especie en los últimos miles de años.
Introducción
entre el conocimiento y el pensamiento científico. Algunas sobrepasan este ámbito y se convierten en el centro de las grandes preguntas que se hace la humanidad. Quiénes somos es el objeto que queremos abordar desde el punto de vista biológico, etológico, social y cultural. Es muy interesante verlo desde una perspectiva holística que nos proporcione una visión completa no solo del proceso de hominización sino también del proceso de humanización.
Por lo que respecta a la dialéctica hominización-humanización, los debates siempre se han movido en la dicotomía entre evolución biológica y evolución cultural. Estos procesos ahora se analizan de otra manera y se llaman «procesos de coevolución». Nosotros preferimos el concepto de evolución integrada o proceso convergente. Es verdad que una y otra forma de evolución tienen diferentes ritmos de integración. Mientras que durante las primeras fases la progresión biológica y la cultural eran lentas, durante el último millón de años estas se han ido acelerando, hasta llegar al punto actual, en el que nuestra especie se encuentra en proceso de evolución cultural y técnica exponencial, sobre todo en los últimos decenios.
«En la dialéctica hominización-humanización, los debates siempre se han movido en la dicotomía entre evolución biológica y evolución cultural»
Ahora, mientras se producen pocas modificaciones de tipo biológico, se están desencadenando de forma acelerada muchas adquisiciones culturales. Llamamos «complejidad» al conjunto de adquisiciones que nos permiten diferenciarnos específicamente de los otros animales. Entre las biológicas, nos referimos principalmente al crecimiento del encéfalo y, por lo que respecta a las culturales y técnicas, que son diversas y en muchos casos secuenciales, estamos hablando de tecnología lítica, del fuego, del lenguaje o bien de enterrar a los muertos o del simbolismo, entre otras adquisiciones.
Los humanos actuales nos definimos por habernos convertido en seres complejos. La cuestión es si la complejidad es exclusiva de nuestra especie o si ha emergido y se ha consolidado en otras especies (Agustí y Carbonell, 2013).
Solamente contestando estas preguntas, a partir de datos analíticos, seremos capaces de responder la cuestión de quiénes somos y si somos fruto de las adquisiciones de otras especies; así mismo podremos definir, cuando existan, los rasgos específicos de nuestra especie respecto de las otras de nuestro género.
Nos referimos a quiénes somos los Homo sapiens. Tenemos que empezar buscando nuestro origen en el espacio y nuestra posición filogenética. Después podremos referirnos a las adquisiciones. Eso quiere decir hablar de dónde venimos, conocer nuestras raíces o, por lo menos, plantear una hipótesis sobre esta cuestión.
Rastreando el origen
Los restos más antiguos de nuestra especie se encuentran en África. Son los de Kibish, en Etiopía, representados por Omo 1 y Omo 2, que tienen una cronología de 195.000 años. En el momento de ser descubiertos en 1967, se pensaba que eran menos antiguos, pero unas dataciones posteriores dieron la sorpresa: son 40.000 años más viejos que los restos de Herto (Shea et al., 2002), también de Etiopía, pertenecientes todos ellos a la subespecie Homo sapiens idaltu (White et al., 2003).
Hace 100.000 años, Homo sapiens ya se encontraba diseminado por toda África. En Sudáfrica es representativo el yacimiento de Klasies River y, en el norte del continente, el de Jebel Irhoud (Marruecos), con una antigüedad semejante a Herto. A finales del Pleistoceno medio, hace unos 120.000 años, Homo sapiens partió del continente africano y lo encontramos desperdigándose por Asia y llegando después a Australasia hace unos 60.000 años (Gillespie, 2002). Finalmente, llegó a Europa hace unos 45.000 años, antes de ocupar la última frontera: el continente americano.
El proceso de ocupación del planeta ha continuado hasta hoy, cuando nuestra especie ha pisado toda la superficie terrestre y la ha ocupado de forma continua excepto pequeñas discontinuidades.
¿Cuáles son nuestros orígenes biológicos? Esta es otra cuestión interesante que aún nos es desconocida pero que ha dado pie a algunas hipótesis. La lista de los homínidos que precedieron a Homo sapiens en África incluye a Homo rodhesiensis, cuyos fósiles más característicos son, entre otros, los de Broken Hill (Zambia) y Bodo (Etiopía).
Tras encontrar a Homo antecessor, el equipo del yacimiento español de Atapuerca (Burgos) publicó en 1997 que las características craneales de este hominino lo convertían en candidato a ser el origen común de neandertales en Europa y Homo sapiens en África (Bermúdez de Castro et al., 1997).
Esta hipótesis tan arriesgada nos viene dada por las características tan modernas del maxilar, en el que destacan la falta de prognatismo y las depresiones suborbitales marcadas, que le dan una morfología igual a la de nuestra especie. También porque H. antecessor tiene caracteres primitivos del clado Homo, caracteres compartidos con otros homininos del Pleistoceno medio, así como un conjunto de rasgos compartidos con los neandertales (Bermúdez de Castro et al., 2017). Estos caracteres que ahora nos distinguen de otras especies paleontológicas son los que ya emergieron hace cerca de un millón de años.
Se trata, sin embargo, de definir cuáles son las características de Homo sapiens con respecto a nuestros coetáneos que se extinguieron hace unos 30.000 años. Entre 70.000 y 30.000 años fue el período en que convivieron más especies en nuestro género: los últimos Homo erectus, los denisovanos, Homo neanderthalensis y Homo floresiensis.
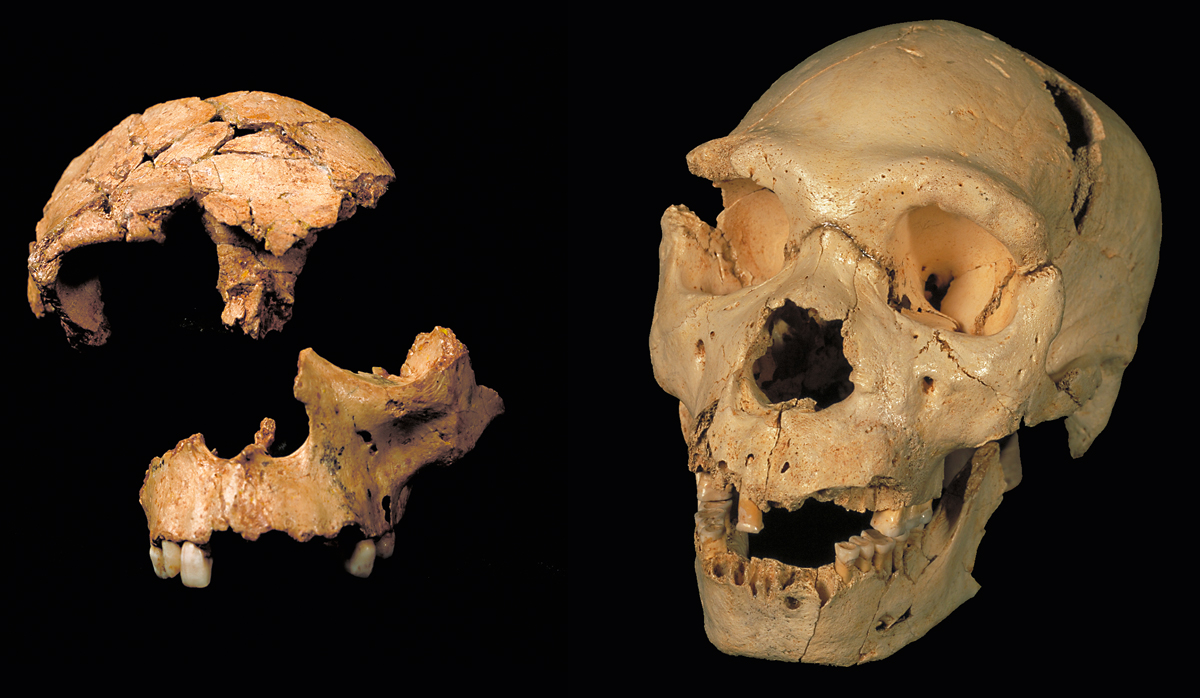
Izquierda: Composición de los restos ATD6-15 (fragmento de cráneo) y ATD6-69 (maxilar), que podrían pertenecer al mismo individuo de Homo antecessor. El conjunto se conoce como «el chico de la Gran Dolina». H. antecessor tiene caracteres primitivos del clade Homo, compartidos con otros homininos del Pleistoceno medio, así como un conjunto de rasgos que comparte con los neandertales. Derecha: Cráneo número 5 del yacimiento de la Sima de los Huesos, en la Cueva Mayor de la sierra de Atapuerca. Los hallazgos de este yacimiento han permitido conocer que los huesos del oído de los humanos de la Sima de los Huesos estaban preparados para recibir en la banda intermedia y ancha, tal como ocurre en los humanos actuales. Eso quiere decir que estaban preparados para capturar un lenguaje que vehiculara sus acciones. / Equipo del Museo Nacional de Ciencias Naturales, CSIC
Características anatómicas y biológicas ‘sapiens’
Homo sapiens se caracteriza anatómicamente por los siguientes rasgos: un esqueleto craneal redondeado y una frente erguida, que presenta un gran volumen encefálico, con una media de 1.350 cm3; una cara plana y más reducida, con las placas infraorbitales orientadas coronalmente; presencia de mentón en la mandíbula; una dentición caracterizada por una reducción con respecto a la de especies anteriores; y, para acabar, el esqueleto poscraneal es más grácil que en otras especies de su género.
Estructura genética e hibridación
Durante muchos años se había pensado que la especie paleontológica Homo sapiens era también una especie biológica1. Algunas pruebas, como las que se encontraron en Lagar Velho (Portugal), nos pusieron sobre la pista de la existencia de caracteres híbridos en un recién nacido, información que no fue admitida por muchos de los colegas paleoantropólogos cuando se presentó el descubrimiento.
La evolución de los métodos de secuenciación finalmente ha permitido comprobar que algunas poblaciones de Homo neanderthalensis se cruzaron con humanos anatómicamente modernos y que en la actualidad los pobladores eurasiáticos tenemos un pequeño porcentaje de genes de los neandertales. Por su parte, también en Oceanía los humanos actuales de la región retienen un pequeño polo genético arcaico, pero en este caso corresponde a genes de los denisovanos, una población ancestral descubierta en una cueva de Siberia y descrita únicamente mediante análisis genético (Krause et al., 2010).
«No solo tenemos antecesores comunes con otras especies, sino que, además, nos hemos hibridado con especies paleontológicas que han convivido con nosotros»
Una vez conocido parte del genoma de estas especies y su historia genética, hay que plantear que probablemente solo se trata de especies paleontológicas y no de especies biológicas. En el sentido estricto de la palabra, se trataría de variabilidad biológica. Por tanto, ni tan siquiera en este terreno los humanos anatómicamente modernos seríamos específicamente diferentes. No solo tenemos antecesores comunes con otras especies sino que, además, nos hemos hibridado con especies paleontológicas que han convivido con nosotros durante los 200.000 años de nuestra evolución. Ahora en nuestro planeta solo existen Homo sapiens, pero eso solo ocurre desde hace unos 30.000 años. Aun así, como hemos dicho, somos probablemente una síntesis filogenética.
El lenguaje
Para que podamos hablar ha sido necesario que se produjesen una serie de modificaciones genéticas, anatómicas y neuronales. Por lo que respecta a las transformaciones en la anatomía, estas modificaciones van desde el desarrollo del área frontoparietal hasta los cambios en el tracto vocal. Ahora sabemos que eso está presente en diferentes especies de nuestro género; por lo menos en algunas de estas ya lo hemos podido contrastar.
Ahora mismo tenemos una serie de pruebas indirectas que nos hacen pensar que el lenguaje puede ser una característica del género Homo y no de la especie Homo sapiens, como formulaba Noam Chomsky (1985). Podría ser innato en el género Homo y, por tanto, tampoco sería una especificidad de nuestra especie, como antes se había dicho. Al menos sabemos que los homininos de hace cerca de 500.000 años podían comunicarse verbalmente con fonemas y morfemas como lo hacemos nosotros. La prueba se ha conseguido gracias al análisis de fósiles obtenidos en la Sima de los Huesos en Atapuerca (Martínez et al., 2004).
Los huesos del oído medio nos han servido para dar respuesta a la pregunta de si la recepción sonora humana semejante a la actual es o no es antigua. El estudio de los fósiles de la Sima de los Huesos nos dice que los humanos que la habitaban podían recibir en la banda intermedia y ancha como nos pasa a los humanos actuales, lo que quiere decir que tendría que haber emisarios que capturaban su lenguaje, es decir, una lengua que permitía vehicular sus acciones no solo en lenguaje corporal como hacen los simios o con interjecciones o expresiones más o menos complejas, sino con la palabra. La estructura y morfología del oído medio indican que los homínidos de la Sima escuchaban con banda ancha, mientras que los otros simios, no; y tampoco tienen habla.
Por lo que respecta a la genética, la presencia del gen FOXP2 (Krause et al., 2007), tanto en Homo neanderthalensis como en Homo sapiens, nos indica que las dos especies tenían el lenguaje fijado hace decenas de miles de años. En los fósiles de Homo neanderthalensis de la cueva de El Sidrón (Asturias) se ha podido identificar este gen relacionado con el lenguaje. Por tanto, esta tampoco es una característica específica de Homo sapiens, sino que, como ya hemos dicho, la comparte el género Homo (Whitfield, 2008).

Vista de los yacimientos de la Trinchera del Ferrocarril de la sierra de Atapuerca, entre los que está la cueva de la Gran Dolina, donde se encontraron los restos de Homo antecessor. El equipo de Atapuerca publicó en 1997 que las características craneales de este hominino lo hacían candidato a ser el origen común de neandertales en Europa y de Homo sapiens. / Jordi Mestre / IPHES
Características culturales y técnicas
Existe una cuestión importante que nos fuerza a distinguir entre biología y cultura. Aunque no se han producido grandes cambios en el esqueleto desde la aparición de los humanos anatómicamente modernos, sí que se han dado grandes cambios culturales desde la aparición de las características anatómicas que nuestra especie adquirió por evolución cultural.
«Algunas de las actividades consideradas complejas no son solamente de nuestro género, pero es nuestra especie la que acaba generalizándolas a lo largo de su evolución»
Algunas de las actividades consideradas complejas desde una perspectiva actual no son propias solamente de nuestro género, pero sí que podemos decir que es nuestra especie la que acaba generalizándolas a lo largo de su evolución. Las detallamos a continuación.
Homo sapiens se caracteriza culturalmente y socialmente por varias cuestiones. En el campo tecnológico, las industrias líticas experimentan una gran evolución en el transcurso del proceso de adaptación de nuestra especie que se desarrolla en África y también en su salida de aquel continente. Mientras que las primeras tecnologías se incluyen en la Edad Media de Piedra (Middle Stone Age o MSA), las herramientas caracterizadas por la transición del Modo 2 al Modo 3 y el Modo 42 muestran una síntesis y evolución del sustrato con una gran diversificación, desde la presencia masiva de la técnica Levallois (preparación de núcleos de piedra antes de la extracción de lascas) hasta la utilización sistemática del hueso como materia prima, así como la presencia de la técnica laminar y, al final, la reducción microlítica. La producción de láminas no la emplea solo nuestra especie; otras especies, como los neandertales, también la desarrollaron en el transcurso de su evolución cultural.

Bifaz de cuarcita, bautizado con el nombre de Excalibur, recuperado en el yacimiento de la Sima de los Huesos de la Cueva Mayor de la sierra de Atapuerca. La transición desde el uso de estas herramientas hacia lascas y láminas demuestra que existe una síntesis y evolución con una gran diversificación. La producción de láminas no solo es propia de nuestra especie, otras como los neandertales también la desarrollaron. / Susana Sarmiento
Lo que podemos decir es que, como pasa con otras adquisiciones, Homo sapiens acaba generalizando su producción en todas las poblaciones a escala planetaria, pero comparte estos conocimientos con otras especies. Por lo que respecta a las herramientas de madera, se desarrollaron desde muy antiguo; las lanzas y otros elementos vegetales encontrados en Schöningen (Alemania), con una cronología de más de 300.000 años, así lo indican (Thieme, 1997).
Si nos referimos a las herramientas de hueso, aunque los neandertales también las fabricaron, las secuencias de arpones del Pleistoceno superior de Zambia, de 60.000 años, ya nos indican una fuerte complejidad de estos homininos, una complejidad diferente de la que conocemos en otras especies de esta cronología.
La construcción de cabañas
La construcción con materiales perecederos es muy antigua. Tampoco aquí Homo sapiens es único. Desde las antiguas posibles cabañas de Olduvai (Tanzania) de hace 1,9 millones de años hasta las cabañas de Terra Amata (Francia), o bien las de los neandertales, todas las especies han construido lugares de refugio. Por tanto, la arquitectura tiene un largo recorrido de género y no solo de especie. Si bien es cierto que las cabañas gravetienses construidas con huesos de mamuts al este de Europa presentan una gran complejidad, superior a cualquier otra anterior, lo que indican precisamente es esta larga tradición de construcción entre los humanos.
El fuego y los hogares
El descubrimiento del fuego y el uso social que se le ha dado es muy antiguo. Tenemos evidencias empíricas de la cueva de Wonderwerk en Sudáfrica, donde poblaciones de Homo erectus ya lo utilizaban para iluminarse, calentarse y cocinar hace cerca de un millón de años. El fuego tardará en socializarse más de 600.000 años (James et al., 1989). Los neandertales utilizaron el fuego de forma sistematizada igual que lo hace Homo sapiens. Podríamos decir que cuesta encontrar diferencias entre lo que hicieron unos y otros con una gran variabilidad de medidas y formas: hogares planos, hogares en covacha, hornos linterna, construidos con piedra, en el interior de gours (“hoyas”), en abrigos naturales, etc. Eso también demuestra que la complejidad no emerge solo en nuestra especie y que la generalización del fuego es inespecífica. Representa una larga tradición adaptativa en nuestro género que acaba convirtiéndose en un rasgo esencial para las especies que la introducen como mecanismo de socialización.
El fuego y su estructura, el hogar, es muy importante a la hora de continuar desarrollando el lenguaje y sobre todo el lenguaje articulado. La radialización permite la relación intergeneracional y el paso de información, además de todas las funciones primitivas, como dar luz para alargar las horas de actividad, espantar a los animales, perfeccionar herramientas o cocinar.
Ritual funerario
Probablemente la Sima de los Huesos es el primer lugar donde empíricamente se puede probar la existencia de una acumulación intencionada de cadáveres de la historia evolutiva de nuestro género: 28 individuos fueron arrojados al fondo de una sima de 12 metros de profundidad hace unos 400.000 años (Arsuaga, Martínez, Gracia, Carretero y Carbonell, 1993; Carbonell et al., 2003). Eran individuos de la especie Homo heildebergensis. Más tarde, Homo neanderthalensis inició también el comportamiento de enterrar a sus muertos. Tenemos pruebas de este comportamiento neandertal desde Europa hasta el próximo Oriente, aunque algunos colegas no están de acuerdo (Pettitt, 2002). Los ejemplos incluyen yacimientos como Shanidar 4 en Irak, La Ferrassie en Francia o Kebara en Israel. Por tanto, los rituales funerarios no son una adquisición solo de los humanos modernos, sino que este hábito lo comparten otras especies.
Arte y música
El arte es una expresión simbólica básica de nuestra especie. Eso era por lo menos lo que pensábamos hasta hace poco. Ahora tenemos pruebas de actividad artística en el Pleistoceno medio, en dos yacimientos aixelianos. Nos referimos a la Venus de Tam Tam, encontrada en Marruecos con 400.000 años de antigüedad, y la Venus de Berekhat Ramo, en los Altos del Golán, de más de 200.000. Se trata de esculturas, una producción artística en tres dimensiones (D’Errico y Nowell, 2000).
También disponemos de una serie de grabados intencionados en el yacimiento del Pleistoceno medio de Bilzingsleben, en Alemania. Podemos decir, pues, que, con una alta probabilidad, el arte no es un invento específico de Homo sapiens, sino que lo es también de otras especies, algunas ellas contemporáneas y otras que nos han precedido.
Lo que sí que podemos afirmar es que el arte lo ha generalizado nuestra especie desde hace unos 45.000 años, pero no antes. Aunque haya testimonios anteriores, hasta esta fecha no se hace masivo y se extiende por nuestro planeta. Es con esta cronología que encontramos instrumentos de viento como las flautas. A partir de este momento, la presencia de esta actividad simbólica es universal y, por tanto, la encontramos en todos los continentes.
Conclusión
Lo hemos planteado en otras ocasiones y aquí también lo hemos dejado caer: los humanos anatómicamente modernos representamos una síntesis evolutiva, biológica y cultural de nuestro género. La genética nos ha permitido contrastar la existencia de multitud de hibridaciones de las poblaciones que vivieron y convivieron con Homo sapiens fuera de África.
«‘Homo sapiens’ no es la especie elegida ni la culminación de nada, sino que representa un proceso normal, aunque singular, de la selección natural»
Si técnica y culturalmente ya se habían apreciado analogías en las industrias líticas de diferentes especies, ahora podemos concluir que este hecho converge con los resultados de las pruebas genéticas obtenidas en registros fósiles. Estas diferentes poblaciones no tan solo tuvieron contactos culturales, sino que debieron de mezclarse en varias ocasiones y establecer mecanismos biológicos y culturales para pasarse información. En realidad tenemos que hablar, tal como ya hacemos, de evolución integrada y compartida.
Homo sapiens, contrariamente a lo que algunos pueden pensar, no es la especie elegida ni la culminación de nada, sino que representa un proceso normal, aunque singular, de la selección natural. Lo que pasa es que el aumento exponencial de complejidad de la especie, sobre todo por lo que respecta al comportamiento cultural y técnico (herramientas, lenguaje, fuego, arte, entierro de los muertos, etc.) nos ha permitido incrementar la sociabilidad de manera inconmensurable, como no había pasado nunca en ninguna otra población de seres vivos en el planeta. La capacidad de socialización que han proporcionado las adquisiciones realizadas por nuestra especie es la responsable de la emergencia de una conciencia compleja. Es responsable de que seamos como somos.
- Una especie biológica es aquella en la que los individuos masculinos y los femeninos tienen descendencia fértil después de cruzarse. Por otra parte, unos restos esqueléticos pertenecen a una especie paleontológica cuando tienen una serie de características morfológicas asociadas que los diferencian de otras asociaciones esqueléticas. (Volver al texto)
- Mientras que el Modo 2 se caracteriza por las industrias de gran formato como bifaces, picos o hendedores, en el Modo 3 destacan las lascas y las lascas retocadas, y en el Modo 4, las láminas y lascas muy estilizadas. (Volver al texto)
REFERENCIAS
Agustí, J., & Carbonell, E. (2013). L’evolució sense sentit. Barcelona: Empúries.
Arsuaga, J. L., Martínez, I., Gracia, A., Carretero, J. M., & Carbonell, E. (1993). Three new human skulls from the Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain. Nature, 362, 534–537. doi: 10.1038/362534a0
Bermúdez de Castro, J. M., Arsuaga, J. L., Carbonell, E., Rosas, A., Martínez, I., & Mosquera, M. (1997). A hominid from the Lower Pleistocene of Atapuerca, Spain: Possible ancestor to Neanderthals and modern humans. Science, 276(5317), 1392–1395. doi: 10.1126/science.276.5317.1392
Bermúdez de Castro, J. M., Martinón-Torres, M., Martín-Francés, L., Modesto-Mata, M., Martínez de Pinillos, M., García, C., & Carbonell, E. (2017). Homo antecessor: The state of the art eighteen years later. Quaternary International, 433, 22–31. doi: 10.1016/j.quaint.2015.03.049
Carbonell, E., Mosquera, M., Ollé, A., Rodríguez, X. P., Sala, R., Vergès, J. M., … Bermúdez de Castro, J. M. (2003). Les premiers comportements funéraires auraient-ils pris place à Atapuerca, il y a 350 000 ans? L’Anthropologie, 107(1), 1–14. doi: 10.1016/S0003-5521(03)00002-5
Chomsky, N. (1985). El conocimiento del lenguaje. Madrid: Alianza Editorial.
D’Errico, F., & Nowell, A. (2000). A new look at the Berekhat Ram figurine: Implications for the origins of symbolism. Cambridge Archaeological Journal, 10, 123–167. doi: 10.1017/S0959774300000056
Gillespie, R. (2002). Dating the first Australians. Radiocarbon, 44, 455–472. doi: 10.1017/S0033822200031830
James, S., Dennell, R., Gilbert, A., Lewis, H., Gowlett, J., Lynch, T., ... James, S. (1989). Hominid use of fire in the Lower and Middle Pleistocene: A review of the evidence [and comments and replies]. Current Anthropology, 30(1), 1–26.
Krause, J., Fu, Q., Good, J. M., Viola, B., Shunkov, M. V., Derevianko, A. P., & Pääbo, S. (2010). The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature, 464, 894–897. doi: 10.1038/nature08976
Krause, J., Lalueza-Fox, C., Orlando, L., Enard, W., Green, R. E., Burbano, H. A., … Pääbo, S. (2007). The derived FOXP2 variant of modern humans was shared with neandertals. Current Biology, 17(21), 1908–1912. doi: 10.1016/j.cub.2007.10.008
Martínez, I., Rosa, M., Arsuaga, J.-L., Jarabo, P., Quam, R., Lorenzo, C., … Carbonell, E. (2004). Auditory capacities in Middle Pleistocene humans from the Sierra de Atapuerca in Spain. Proceedings of the National Academy of Sciences, 101(27), 9976–9981. doi: 10.1073/pnas.0403595101
Pettitt, P. B. (2002). The Neanderthal dead: Exploring mortuary variability in Middle Paleolithic Eurasia. Before Farming, 1(4), 1–17. doi: 10.3828/bfarm.2002.1.4
Shea, J. J., Fleagle, J. G., Brown, F. W., Assefa, Z., Feibel, C. & McDougall, I. (2002). Archaeological reconnaissance of the Omo Kibish Formation, Ethiopia. Journal of Human Evolution, 42, A33–A34.
Thieme, E. (1997). Lower Paleolithic hunting spears from Germany. Nature, 385, 807–810. doi: 10.1038/385807a0
White, T. D., Asfaw, B., DeGusta, D., Gilbert, H., Richards, G. D., Gary. D., … Clark Howell, F. (2003). Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 742–747. doi: 10.1038/nature01669
Whitfield, J. (2008). Evolución. El gen del “lenguaje”, FOXP2, parece esencial para la vocalización animal. Investigación y Ciencia, 379, 10–11.




